ПВХ мембрана LOGICROOF V-SR, 1,5 мм (1,0*10 м), серый, 2 рулона (20 кв.м) (ПВХ и ТПО мембраны)
ПВХ мембрана LOGICROOF V-SR, 1,5 мм (1,0*10 м), серый, 2 рулона (20 кв.м)
Кровельные гидроизоляционные неармированные ПВХ мембраны премиум-класса
Кровельные гидроизоляционные неармированные ПВХ мембраны премиум-класса
Читать все Скрыть- Страна происхождения
- Россия
- Водопоглощение по объему, % не более
- 0,2
- Температурный режим использования
- от -20°С до +50°С
- Армирование
- Нет
- Горючесть, степень
- Г4
- Верхняя сторона
- Гладкая
- Размер
- 1,0 х 10 м
- Цвет
- серый
- Толщина, мм
- 1,5
- Температура гибкости на брусе R=25 мм, не выше, С
- -40
-
Доставка
на следующий день после оплаты
-
Безопасность платежа
технология 3D Secure для карт VISA и Mastercard Secure Code
-
Гарантия качества
прямая покупка от производителя
Одноклассники
Вконтакте
- Показатель
- Значение
- Страна происхождения
- Россия
- Способ монтажа
- Механическая фиксация
- Водопоглощение по объему, % не более
- 0,2
- Температурный режим использования
- от -20°С до +50°С
- Армирование
- Нет
- Горючесть, степень
- Г4
- Верхняя сторона
- Гладкая
- Размер
- 1,0 х 10 м
- Гарантийный срок хранения, месяцев
- 18
- Вес материала
- 2 кг/кв.


- Вид конструкции
- Кровля
- Сопротивление динамическому продавливанию (ударная стойкость), мм, не менее, (по твердому основанию / по мягкому основанию)
- 700/1000
- Сопротивление динамическому продавливанию (ударная стойкость), С, не должно быть трещин
- -30
- Температура гибкости на брусе R=25 мм, не выше, С
- -40
- Полная складываемость на фальце, С, не должно быть трещин
- -30
- Удлинение при максимальной нагрузке, %, не менее
- 19
- Прочность при растяжении, метод А, Н/50 мм, не менее, вдоль рулона / поперек рулона
Применяются для усиления гидроизоляционного ковра в углах кровели, герметизации сопряжений кровли с трубами, антенными растяжками, опорами под оборудования и т.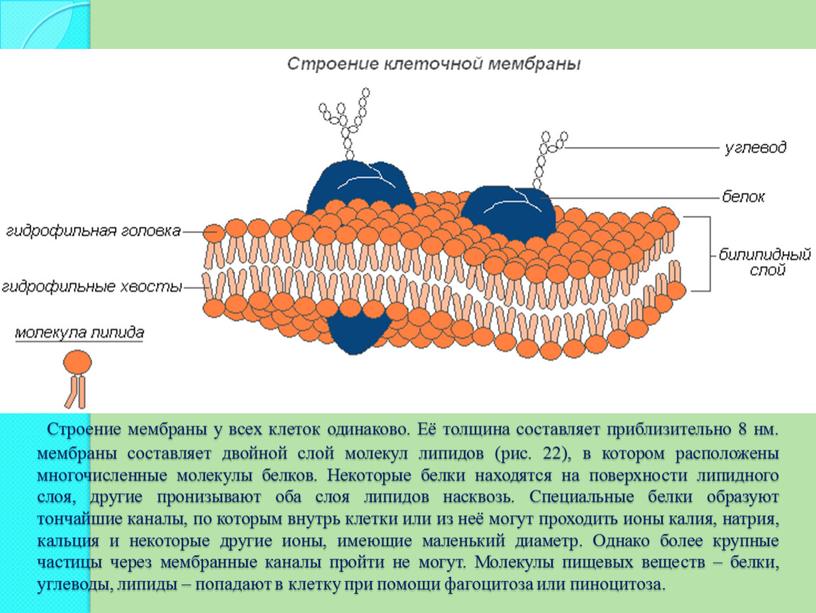 д. Легко растягивается после прогрева горячим воздухом.
д. Легко растягивается после прогрева горячим воздухом.
Производство LOGICROOF
Укладка полимерных мембран.
Укладка полимерных мембран. Основные принципы.
Монтаж мембраны LOGICROOF часть1
Монтаж мембраны LOGICROOF часть2
ПВХ мембрана LOGICROOF V-SR, 1,5 мм (1,0*10 м), серый, 2 рулона (20 кв.
 м)
м)Отзывы пользователей 1 item(s)
5100
- Брал для гидроизоляции сложной кровли. Мембрана достаточно плотная. Ложится легко и ровно, никаких заломов и помятостей. Если укладывать в хороший нахлест и скреплять специальной лентой, то изоляция отличная.
string(1) "5"
Отзыв пользователя Алексей Мехряков / (Отзыв написан 13.09.2018)
Отзывы пользователей 1 item(s)
5100
- Брал для гидроизоляции сложной кровли. Мембрана достаточно плотная. Ложится легко и ровно, никаких заломов и помятостей. Если укладывать в хороший нахлест и скреплять специальной лентой, то изоляция отличная.
string(1) "5"
Отзыв пользователя Алексей Мехряков / (Отзыв написан 13.09.2018)
 Мембрана достаточно плотная. Ложится легко и ровно, никаких заломов и помятостей. Если укладывать в хороший нахлест и скреплять специальной лентой, то изоляция отличная.
Мембрана достаточно плотная. Ложится легко и ровно, никаких заломов и помятостей. Если укладывать в хороший нахлест и скреплять специальной лентой, то изоляция отличная.Мембраны Gore-Tex. Что это и как работает?
Н
аверняка вы видели эти яркие этикетки с надписью Gore-Tex на некоторых товарах в наших магазинах. И, может быть, даже задавались вопросом, почему одежда с ними стоит чуть дороже одежды без них. Мы посвятили мембранам Gore-Tex подробный текст, чтобы рассказать обо всех преимуществах технологии и ее применении.
Производители обещают, что в экипировке с мембраной Gore Tex вам в любую непогоду будет тепло и комфортно (почти как дома в халате)
Что такое Gore-Tex?Gore-Tex — это мембрана, которую активно используют очень многие бренды Outdoor одежды. Вся фишка заключается в том, что Gore-Tex защищает от ветра, останавливает воду и, при этом, позволяет водяному пару выйти за пределы ткани. Проще говоря, Gore-Tex даёт одежде “дышать” и не отсыревать внутри даже во время физических нагрузок. То, что нужно для спорта. .Как это работает?
Влага в мембранной одежде (в том числе, с ламинатом Гортекс) отводится за счет разницы давлений воздуха под одеждой и снаружи. Поэтому, если вы, например, присядете отдохнуть на веранде в кафе, то мембрана не поможет. Она работает только тогда, когда вы двигаетесь.А как именно она работает?
Чтобы вам проще было понять, о чем мы говорим, вы можете представить себе изгородь, отражающую порывы ураганного ветра.
 Мембрана изготовлена из расширенного тефлона, растяжение которого образует микропористый материал с 1,7 миллионами пор на квадратный сантиметр. Разными способами эта мембрана интегрируется в ткань одежды, чтобы сделать её полностью водонепроницаемой и ветрозащитной.
Мембрана изготовлена из расширенного тефлона, растяжение которого образует микропористый материал с 1,7 миллионами пор на квадратный сантиметр. Разными способами эта мембрана интегрируется в ткань одежды, чтобы сделать её полностью водонепроницаемой и ветрозащитной.Основные свойства: водостойкость (защита от дождя и мокрого снега), защита от ветра и паропроницаемость (“дышащая” способность ткани)
.
То есть, ткань совсем не промокает?
Нет, не промокает. Мембрана склеена с наружным слоем ткани. Кроме того, лицевая ткань изделия покрывается специальным составом с полимерами. Он не оставляет воде шансов. Особенно, если остальные слои одежды подобраны правильно.Что за слои?
Мы имеем в виду термобелье (базовый слой) и толстовки из флиса или полартека (средний слой).
Каждый слой выполняет свою функцию. В то время, как мембранная одежда защищает от воды и ветра, термобелье впитывает и отводит пот, чтобы защитить организм от перегрева и переохлаждения в процессе нагрузки.
 Ну а флис сохраняет тепло.И тогда холодно не будет?
Ну а флис сохраняет тепло.И тогда холодно не будет? Будет как в той рекламе — сухо и комфортно. Хотя некоторые говорят, что влага иногда все-таки ощущается. Дело в том, что горнолыжная одежда не проветривается так, как обычная майка. И когда вы находитесь без движения, пар задерживается внутри, так как давления, о котором мы говорили выше, не возникает.
Важно! Мембрана не спасает от холода! Для этого есть утеплитель и второй слой одежды.
А кто и когда придумал Gore-Tex?Gore-Tex обнаружили случайно в 1969 году. Проводя эксперимент с нагретыми стержнями из тефлона, инженер-химик Боб Гор в отчаянии сильно растянул один из этих стержней и, сам того не ожидая, открыл совершенно новую форму полимера, теперь известную как ePTFE (извините нас, за небольшой урок химии). Структура этого самого ePTFE образована миллионами узелков. Они настолько маленькие, что частички воды не могут через них пройти. А вот пар может.
Бобу Гору потребовалось несколько лет, чтобы получить необходимые патенты на свою революционную технологию.
 В конечном итоге, бренд Gore-Tex был представлен на рынке в 1978 году.
В конечном итоге, бренд Gore-Tex был представлен на рынке в 1978 году.И что, с того момента ничего не изменилось?
Изначально мембрана не была на 100% неуязвимой — вода все равно просачивалась сквозь швы. Технологию доработали чуть позже, в 1979 году, когда производители разработали специальный станок и ленту Gore Seam для проклейки. В итоге, швы обработали по принципу швов гидрокостюмов.Как используют Gore-Tex?
Здесь нужно начать с того, что у ламинатов Gore-Tex есть классификация.
Gore-Tex Pro — наиболее прочный и устойчивый к истиранию трехслойный ламинат с самыми высокими показателями водостойкости. Как правило, Gore-Tex Pro используется в экипировке для самых экстримальных погодных условий. Например, для big-mountain фрирайда.
Сноуборд-одежда с мембраной Gore-Tex Pro
Gore-Tex Active проигрывает Pro в плане водостойкости, но не уступает по показателям паропроницаемости.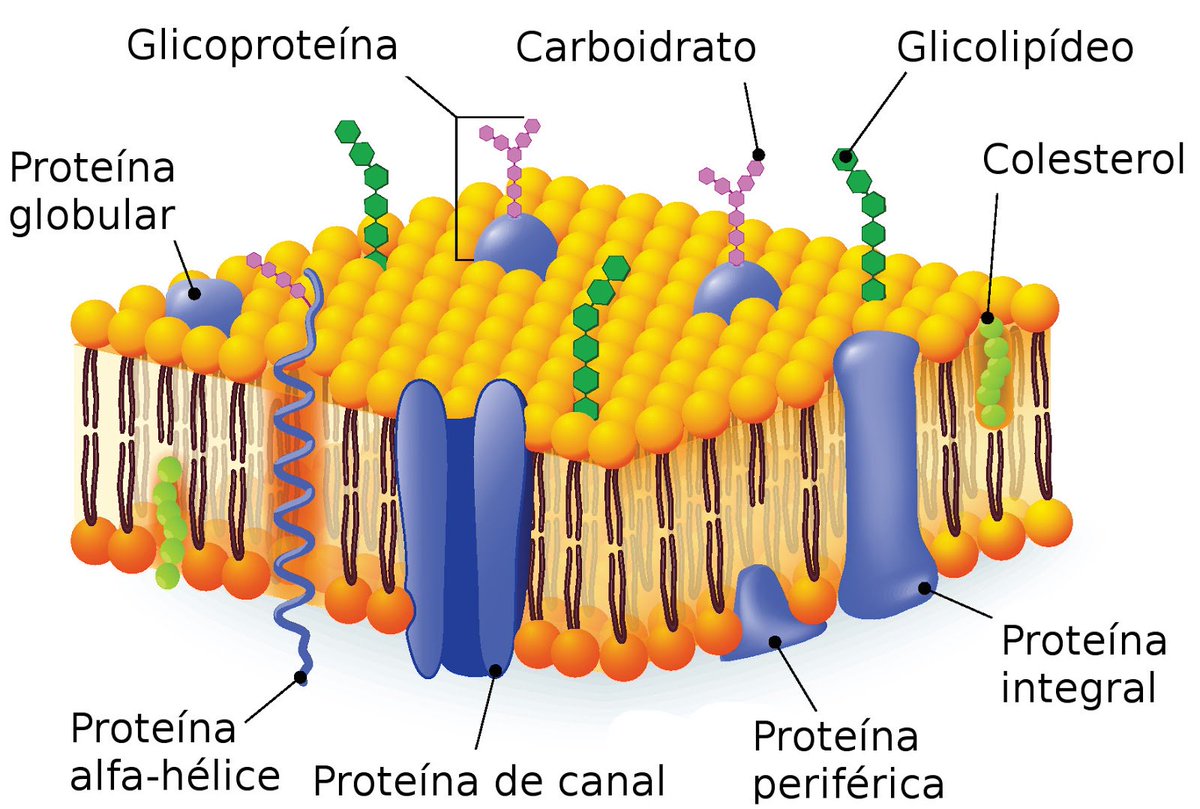 Это максимально “дышащая” одежда, в которой можно хоть куда — она подходит для трекинга, бега, скитура, для катания на сноуборде и лыжах.
Это максимально “дышащая” одежда, в которой можно хоть куда — она подходит для трекинга, бега, скитура, для катания на сноуборде и лыжах.
Сноуборд-одежда с мембраной Gore-Tex Active
Gore-Tex Products — в эту категорию попадает все остальное: кепки, обувь, иногда экипировка для активных видов спорта и повседневная одежда.
Городская одежда с мембраной Gore-Tex
К слову говоря, бренды не сразу поняли весь потенциал Gore-Tex и на первых парах много экспериментировали. Еще до выхода первой коллекции одежды с Gore-Tex в магазинах появились мембранные палатки и спальные мешки. Затем появилась первая парка, которая по крою больше напоминала обычный дождевик. Позже всего свет увидели трекинговые ботинки и перчатки с “дышащей” прослойкой.
Интересный факт: форму с Gore-Tex шьют для военных в США и Британии. Вот так вот… Кому-то кирзачи, а кому-то берцы с мембранной подкладкой.
Что это за цифры в описании одежды Gore-Tex?Это характеристики.
 Они выражаются примерно вот так: 20K/20K или 15 000/10 000, где первые цифры — это показатели водостойкости, а вторые — паропроницаемости. Чем выше числовые значения, тем активнее «работает» мембрана.
Они выражаются примерно вот так: 20K/20K или 15 000/10 000, где первые цифры — это показатели водостойкости, а вторые — паропроницаемости. Чем выше числовые значения, тем активнее «работает» мембрана.Для горнолыжных курток Gore-Tex обычные показатели — это 28K/20K, для курток с другими мембранами эти значения могут опускаться до 10K10K.
Почему одежда Gore-Tex дороже?Как правило, и в России и в Европе средняя цена горнолыжной куртки с Gore-Tex составляет 200-300 долларов. И дело не в себестоимости мембраны (она не так уж высока). Тут математика немного другая. Если бренд хочет делать одежду из Gore-Tex, то он должен соответствовать стандартам качества товарной марки. А именно а) он должен купить разрешение на производство от W. L. Gore & Associates (компании принадлежит патент на технологию). б) этот бренд должен обзавестись сертифицированным оборудованием (в частности, аппаратом для проклейки швов, о котором мы уже писали). Время от времени фабрику инспектируют представители Gore & Associates.
 Они же, кстати, проводят испытания материалов и следят за этапами изготовления.А есть ли недостатки у этих тканей?
Они же, кстати, проводят испытания материалов и следят за этапами изготовления.А есть ли недостатки у этих тканей?В совсем уж лютый холод появляется проблема обмерзания. Пары влаги конденсируются на внешней стороне ткани и она буквально дубеет (а иногда даже шуршит). Бороться с этим невозможно. Да и нужно ли?Как ухаживать за одеждой с Gore-Tex?
Точно также, как и за любой другой мембранной одеждой:
- Не стирать слишком часто, так как защитные свойства ткани могут “смыться” вместе с грязью.
- Обзавестись специальной бытовой химией для мембранных тканей. В крайнем случае, мы можем посоветовать шампуни без хлора — они помогут сохранить целостность ламината и не “забьют” поры ткани.
- Температура для машинной стирки не должна быть выше 40’. Будет идеально, если вы включите еще и “деликатный” режим.
- Не помешает отдельно отправить изделие в сушку минут на 20. Либо просушить одежду самостоятельно при помощи утюга (но только без пара и на низкой температуре!). И не забудьте использовать марлю!
- Специальные пропитки для мембранная одежды — настоящая находка! Есть лубриканты для обработки швов, спреи для защиты от грязи и соли, чистящие пенки и куча других разных фишечек, включая наклейки и заплатки.
 И не забудьте использовать марлю!
И не забудьте использовать марлю!Мембрана Marmot MemBrain® | День сурка
Marmot Membrain™
Marmot Membrain™ использует усовершенствованную технологию, которая сочетает в себе наилучшую эффективность воздухопроницаемости, водостойкости и водоотталкивающих свойств с непревзойденной надежностью. С минимальной эффективностью в 10, 000mm / 10,000mm она гарантированно сохранит вас в сухости и комфорте в наихудших погодных условиях.
Как это работает?
Технология Marmot Membrain™ обеспечивает отличную защиту от непогоды и внутренний комфорт. Гидрофильная природа ламинирования из PU уменьшает внутренний конденсат и расширяет границы комфорта уменьшая эффект озноба, вызываемый переходом конденсата обратно в пар.
Каковы преимущества этой технологии?
Эта 2-хслойная ламинация сочетается с превосходной лицевой тканью, создавая мягкий воздухопроницаемый и дышащий материал с великолепной эффективностью. Благодаря своей непревзойденной воздухопроницаемости и продолжительной способности надежно отталкивать влагу эта усовершенствованная ткань обеспечивает новый уровень эффективности для искателей по-настоящему эффективной защиты. Водонепроницаемость: 10,000 mm minimum JIS-L 1092 Воздухопроводимость: 10,000 gm / 24h minimum JIGS-1099 B1 RET: 6 to 8 Технология: 2-хслойная гидрофильная PU ламинация
Благодаря своей непревзойденной воздухопроницаемости и продолжительной способности надежно отталкивать влагу эта усовершенствованная ткань обеспечивает новый уровень эффективности для искателей по-настоящему эффективной защиты. Водонепроницаемость: 10,000 mm minimum JIS-L 1092 Воздухопроводимость: 10,000 gm / 24h minimum JIGS-1099 B1 RET: 6 to 8 Технология: 2-хслойная гидрофильная PU ламинация
Marmot MemBrain® Strata™
Сверхлегкая, водонепроницаемая/дышащая ткань Marmot MemBrain® Strata™. Показатели мембраны: 20000 мм — влагозащита, 20000 г — дышимость. В отличие от 3-х слойных ламинатов, MemBrain® Strata™ обладает большей паропроводимостью и легким весом. Ткань Strata™ разработана из мелких неорганических частиц смешанных с частицами ткани MemBrain®, которые препятствуют быстрому вытиранию ткани и защищают от дождя. Задняя часть ткани — максимально прочная и паропроводимая. По результатам тестов показатели ткани Strata™ в 4-5 раз выше, чем у обычной MemBrain®. Если сравнивать 2.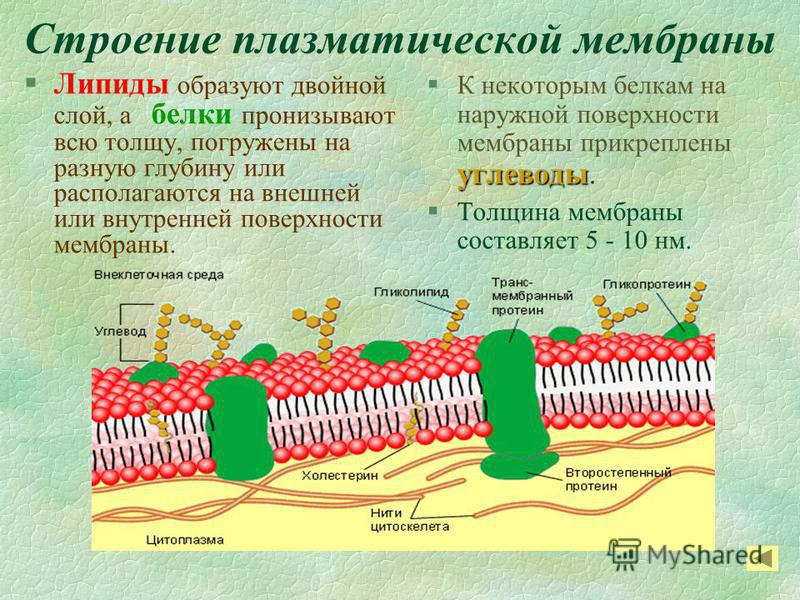 5 — слойную мембрану Marmot с аналогичными тканями других фирм, то Strata™ отличается от них, в первую очередь, — великолепной паропроводимостью (на 100% более дышащая), большей прочностью и, к тому же, она приятна на ощупь.
5 — слойную мембрану Marmot с аналогичными тканями других фирм, то Strata™ отличается от них, в первую очередь, — великолепной паропроводимостью (на 100% более дышащая), большей прочностью и, к тому же, она приятна на ощупь.
Показатели:
Водонепроницаемость: 40+; PSI 20 000mm
Водоотталкивающие свойства: (% к колич. стирок) 80/100
Ret : 2,5 слоя — 6-6,5 JIS L-1099
B1: 2 слоя — >
20 000 г/м/24 часа
Ветронепроницаемость: O CFM (100%)
Marmot MemBrain® FusionDri
Новая эксклюзивная трехслойная мембрана от Marmot, которая обеспечивает непревзойденную защиту от ветра и влаги, дышимость, легкость и комфорт при высоких аэробных активностях. — внешний слой защищает от суровых внешних погодных воздействий — средний слой гарантирует максимальную дышимость и водонепроницаемость (На основе мембраны MemBrain®) — внутренний слой обеспечивает быстрое отведение влаги за счет ультралегких волокон DriClime. Комбинация 3-х слоев позволяет внутренним водяным парам равномерно распределяться по всей поверхности мембраны и очень быстро испаряться.
Показатели:
Водонепроницаемость: 30,000мм
Паропроницаемость: 50,000 г/м/24часа
Ветронепроницаемость: 0 CFM (100%)
Marmot MemBrain® Eco
Marmot Membrain® Eco — это передовая мембранная ткань. Технология этой мембраны гармонично сочетает отличную воздухопроницаемость, водонепроницаемость и экологическую защиту окружающей среды. Membrain® Eco не содержит перфторированные соединения и не использует их при изготовлении или в процессе ламинирования ткани.
Что такое хирургическая мембрана и для чего она нужна?
По мере того, как медицина погружается глубже в освоение проблематики заболеваний, появляются новые методики решения этих проблем; соответственно, для новых методик требуются новые технологии и материалы.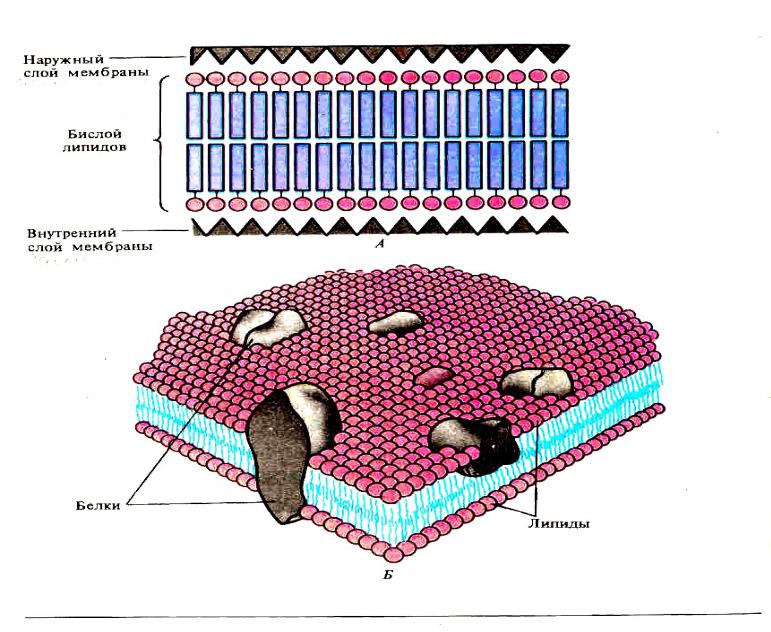 Так появились хирургические мембраны, успешно применяемые в стоматологии.
Так появились хирургические мембраны, успешно применяемые в стоматологии.
Такие мембраны представляют собой тонкие эластичные плёночные фрагменты, применяемые при хирургических/имплантологических операционных вмешательствах. Подробнее рассмотрим, в каких случаях применяются мембраны, что даст более полное представление о том, что это такое.
Наибольшее применение хирургические мембраны нашли в дентальной имплантации.
Напомним, что «классическая» методика проведения имплантации подразумевает два этапа: внедрение имплантата и последующую ортопедическую часть запланированного лечения (то есть, установку протеза на прижившийся имплантат). Так как обычно после утраты зуба и до операции по вживлению искусственного корня проходит довольно большой период, работам непосредственно с имплантатом предшествует операция по реставрации необходимого объёма кости (остеопластика), в связи с тем, что наблюдается атрофия костной ткани.
При проведении таких операций врач использует либо фрагменты костной ткани самого пациента (подсадка костных блоков), либо материалы/препараты, способствующие росту кости.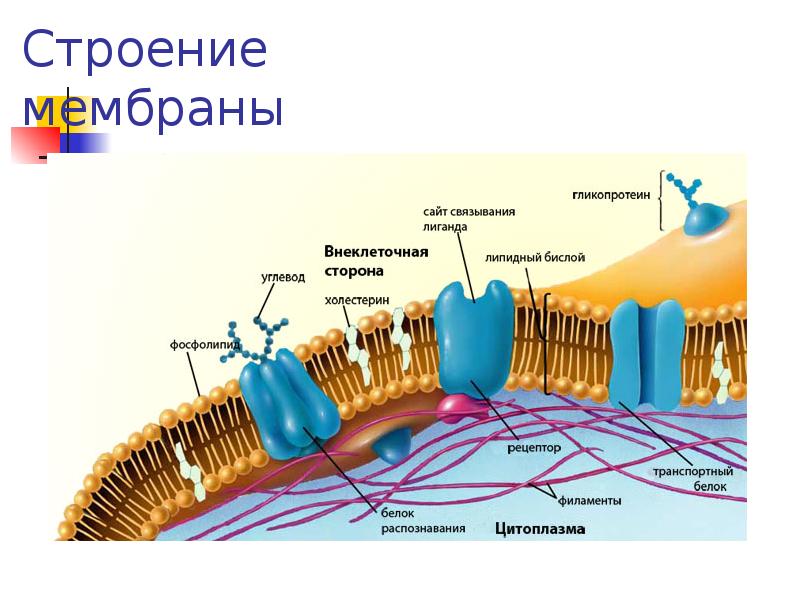 И в том, и в другом случае хирург может применить мембрану (костные блоки могут быть закреплены титановыми винтами; для фиксации остеопластического материала использование мембраны целесообразно и обусловлено физическими свойствами материала). В данном случае мембрана выполняет как бы армирующую функцию: удерживает костнопластический материал в необходимом правильном положении. Как армирующее средство мембрана может быть применена и при имплантации с одномоментной подсадкой остеопластического материала. Мембрана может быть закреплена на кости или на соседних от места вмешательства зубах.
И в том, и в другом случае хирург может применить мембрану (костные блоки могут быть закреплены титановыми винтами; для фиксации остеопластического материала использование мембраны целесообразно и обусловлено физическими свойствами материала). В данном случае мембрана выполняет как бы армирующую функцию: удерживает костнопластический материал в необходимом правильном положении. Как армирующее средство мембрана может быть применена и при имплантации с одномоментной подсадкой остеопластического материала. Мембрана может быть закреплена на кости или на соседних от места вмешательства зубах.
Хирургические мембраны могут быть применены при удалении зуба.
Как было упомянуто выше, потеря зуба постепенно приводит к убыли костной ткани. В частности, кость атрофируется, не получая необходимой нагрузки. Риск атрофии костной ткани появляется и после удаления зуба (удаления корня). Дело в том, что костная ткань и мягкие ткани имеют разную способность и скорость регенерации: дёсенная ткань разрастается и как бы занимает пространство, не давая расти кости, восстановление которой «не поспевает» за ростом мягких тканей. В этом случае мембрана выполняет барьерную функцию, препятствуя такому разрастанию дёсенных тканей (таким образом, хирургические мембраны также называют барьерными мембранами).
В этом случае мембрана выполняет барьерную функцию, препятствуя такому разрастанию дёсенных тканей (таким образом, хирургические мембраны также называют барьерными мембранами).
Использование мембран может быть целесообразно для укрепления подвижных зубов при пародонтологических лоскутных операциях.
В современной стоматологии применяется несколько десятков различных мембран, которые можно разделить на две большие группы:
- Резорбируемые. Такие мембраны постепенно рассасываются.
- Нерезорбируемые. В определённый момент врачу надо будет хирургически удалить установленную ранее мембрану.
Мембрану какого именно типа использовать определяется конкретными клиническими данными и планом лечения.
Как видим, помещение мембраны не является самостоятельной процедурой – это часть показанной по определённому случаю операции.
Хирургические операции различного назначения в сети клиник «Здоровая Улыбка» проводятся опытными квалифицированными специалистами.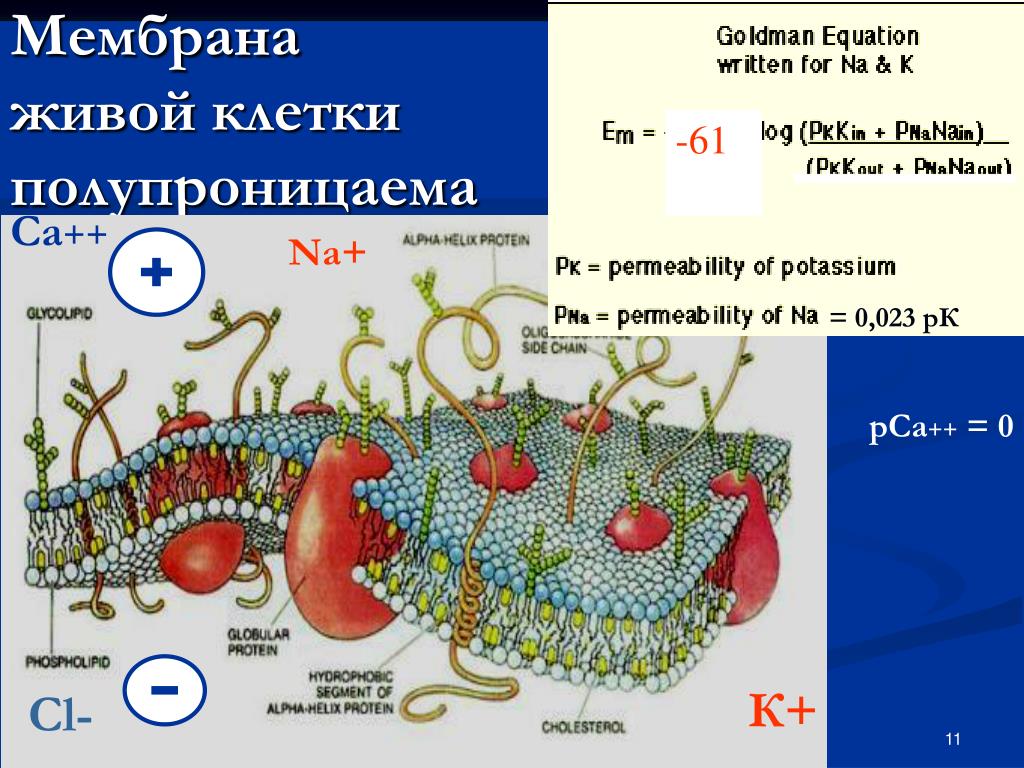
Диффузионная мембрана BRAAS Braas
Описание материала
Трехслойная паропроницаемая мембрана из нетканого полипропилена, с функциональным водонепроницаемым слоем и продольной самоклеящейся полосой на одной стороне полотна. Полоса защищена антиадгезионной полиэтиленовой пленкой, которая удаляется в процессе монтажа.
Область применения
Устройство подкровельного водоизоляционного слоя в конструкциях скатных крыш с рекомендуемыми и малыми уклонами, без сплошного настила.
Способ монтажа
Укладывается по стропилам перпендикулярно направлению ската.
Продольные и поперечные нахлесты полотен должны составлять не менее 100 мм.
Крепится к стропилам в нахлестах скобами степлера.
Герметизация примыканий к вертикальным поверхностям выполняется с помощью
клея Дифодамм тип А.
Преимущества
- обеспечивает свободный вывод влаги из теплоизоляционного слоя;
- обладает стойкостью к механическим повреждениям и ультрафиолетовому излучению.
Технические характеристики
| НАИМЕНОВАНИЕ ПОКАЗАТЕЛЯ, ЕД.ИЗМЕРЕНИЯ | ЗНАЧЕНИЕ |
| Длина, м | 50 |
| Ширина, м | 1,5 |
| Поверхностная плотность, г/кв.м | 120 |
| Масса, кг | 9,9 |
| Эквивалентная толщина слоя воздуха по диффузии пара Sd, м | 0,02 |
| Водоупорность, мм.вод.ст., не менее | 2000 |
| Разрывная сила при растяжении в продольном/поперечном направлении, Н/50 мм, не менее | 220/170 |
| УФ-стабильность, месяцев | 2 |
| Температура эксплуатации, °C | от -40 до +80 |
Упаковка и хранение
| НАИМЕНОВАНИЕ ПОКАЗАТЕЛЯ, ЕД.ИЗМЕРЕНИЯ | ЗНАЧЕНИЕ |
| Количество на поддоне, шт | 36 |
| Размеры поддона, м | 1,2×1,0 |
| Вес поддона, кг | 356 |
Хранить в оригинальной упаковке вертикально в один ярус.
Защищать от влаги и прямых солнечных лучей.
Трехслойная мембрана ИЗОСПАН AM (1600*43750)
Уважаемые покупатели!
ПКФ КроМа предлагает Вам сэкономить время на логистике и доверить нам доставку Ваших заказов, произвденных нашей компанией. Мы имеем свой автопарк и доставляем грузы от 10 кг до 15 тонн, длиной от 0,5 м до 12 метров. В случае необходимости предоставим Вам автомашину с манипуляторм для погрузки и выгрузки тяжелых и габаритных грузов. Мы бережно и в срок доставим Ваш заказ и предоставим полный комплект документов.
- ВНИМАНИЕ ПОКУПАТЕЛЯМ! В соответствии с требованием 54-ФЗ РФ «О применении контрольно-кассовой техники» с 01.07.2019 года вводится обязательный прием оплаты через онлайн-кассы с выдачей контрольно-кассового чека. В связи с этим просим Вас оплачивать доставку при оформлении заказа. Для покупателей не оплативших доставку будет работать дежурная машина с мобильной онлайн-кассой. Спасибо за понимание.

Мы осуществляем доставку:
- В пределах админстративных границ городов: Пенза, Саранск, Кузнецк.
- В населенные пункты Пензенской области и республики Мордовия.
При доставке заказа (товара) Вам необходимо обеспечить наличие подъездных путей по адресу разгрузки для проезда автотранспорта. В случае, если доставка товара осуществляется на закрытую для въезда территорию, Вам необходимо заблаговременно оформить соответствующий пропуск.
Сроки доставки
- Сроки доставки зависят от сроков изготовления заказа.
- Доставка может быть выполнена в день изготовления заказа, либо в другой день (в течении 5 рабочих дней) по желанию клиента.
- Доставка осуществляется в удобный для клиента 2-х часовой интервал, в течении рабочего дня.
Стоимость доставки
Стоимость доставки зависит от веса материала, длины материала, удаленности населенного пункта от склада и от места оформления заказа.
- Скачать тарифы на доставку по г. Пенза:
- Скачать тарифы на доставку по г. Саранск:
- Скачать тарифы на доставку по г. Кузнецк:
- Скачать тарифы на доставку по г. Кузнецк -> область:
- Скачать тарифы на доставку по республике Мордовия:
 Пенза:
Пенза: Разгрузка товара
В стоимость услуги по доставке не входит разгрузка товара. Разгрузка заказанного товара осуществляется силами Покупателя. Для разгрузки тяжелых грузов вы можете заказать автомашину с гидроманимулятором.
Памятка покупателям
При приеме товара внимательно провряйте каждую позицию. В случае выявления дефектов, несоответствия количества или номенклатуры товаров указанным в накладной, Вам необходимо предъявить рекламацию в адрес нашей компании.
Вы можете ознакомиться с подробным переченем гарантийных обязательств на странице «Гарантия».
Наши преимущества
Главные наши преимущества перед другими перевозчиками — пунктуальность и бережное отношение к грузу.
Заказывая у нас доставку, Вы можете быть уверены, что получите свой заказ вовремя и в полной сохранности.
А также:
- Наш транспорт не тентованный, что позволяет осуществлять быструю и безопасную погрузку и разгрузку материала.
- Все наши автомашины оснащены специальными средствами для крепления груза, что исключает любое движение и трение груза по поверхности кузова во время движения.
- Все наши автомашины оборудованы системами глобального навигационного спутникового позиционирования ГЛОНАС, что позволяет координировать маршрут и сроки доставки.
- По требованию Заказчика мы всегда сообщаем точное местоположение автомашины, которая везет его материал, для того чтобы он смог приготовится к приезду груза.
- Системы отслеживания местоположения груза, гарантируют недопущение факта ХИЩЕНИЯ груза.
- Стоимость доставки является фиксированной и не содержит надбавок за дополнительное время.
- Доставка осуществляется в удобное для Заказчика время, место и в любую погоду.
- Мы несем полную ответсвенность за сохранность и качество груза до принятия его покупателем.
- Мы доставляем заказ напрямую строительной бригаде на обьекты Заказичка, без его присутствия.
- Для Вашего удобства Вы можете изменить время и сроки доставки.
- Наши водители всегда готовы помочь Вам с выгрузкой товара.
Как работает мембрана GORE-TEX® – полезная информация от компании Турин
Основные свойства материала обеспечивает мембрана GORE-TEX®, очень тонкая политетрафлуорэтиленовая (PTFE) пленка. Мембрана представляет собой биструктурный микропористый материал, прошедший процесс расширения. Она содержит около 1,4 миллиарда микроскопических пор на один квадратный сантиметр. Каждая пора в 20 000 раз меньше капли воды, но в 700 раз больше молекулы пара. В результате вода в жидком состоянии не проникает сквозь ткань, в то же время пар, выделяемый телом, свободно проходит наружу. Интегрированный в мембрану олеофобный, т. е. жироотталкивающий, компонент создает физический барьер для загрязняющих поры веществ (пот, косметика, пища) и предотвращает тем самым потерю водоотталкивающего и «дышащего» эффектов.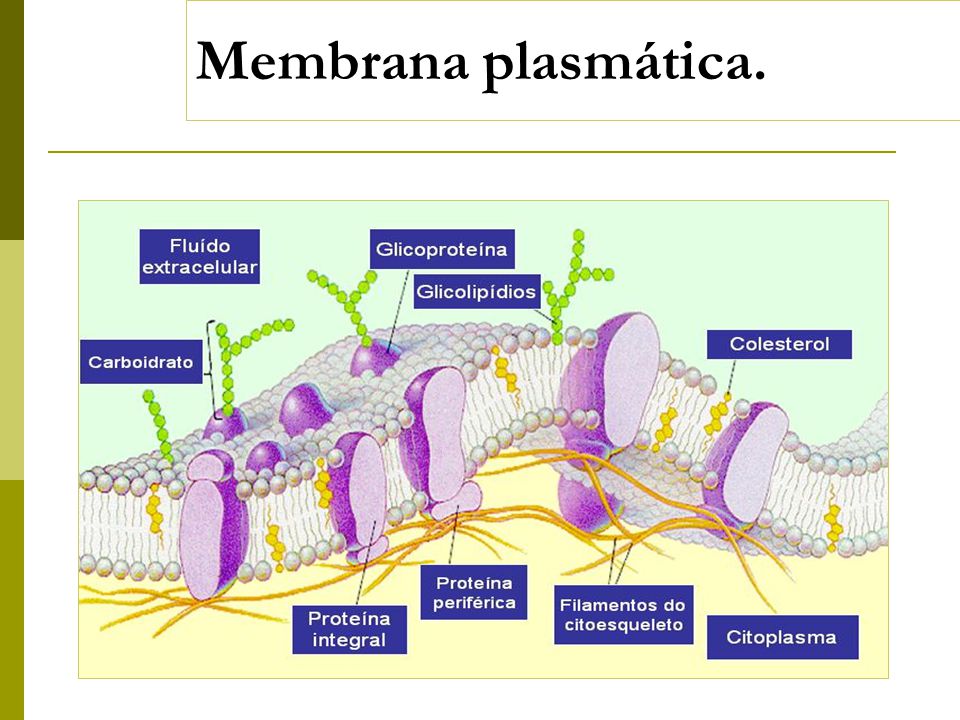
Мембрана GORE-TEX® гарантирует 100% водонепроницаемость.
Водонепроницаемость верхней одежды очень важна во время пребывания на открытом воздухе. Независимо от того, в каких влажных условиях Вы находитесь, ткань GORE-TEX® гарантирует сухость и комфорт.
Мембрана GORE-TEX® «дышит».
Одежда из тканей GORE-TEX® позволяет коже «дышать». Испарения легко проходят через мембрану наружу, обеспечивая естественный процесс терморегуляции и предотвращая возникновение дискомфорта, вызванного перегревом или потерей тепла из-за намокания при интенсивных физических нагрузках.
Мембрана GORE-TEX® гарантирует 100% ветронепродуваемость.
Ткань GORE-TEX® совершенно не пропускает холодный воздух, сохраняя тепло внутри одежды. Благодаря тонковолокнистой структуре мембраны, холодный воздух как бы запутывается в лабиринте микропор, образуя завихрения. Одежда из ткани GORE-TEX® обеспечивает полную защиту от ветра и позволяет дольше оставаться на воздухе при холодной ветреной погоде.
Мембрана GORE-TEX® располагается между внешним материалом изделия и подкладкой. Для производства многослойных материалов GORE-TEX® мембрана прочно соединяется с высококачественными тканями в различных комбинациях. Она создает эффективный барьер для дождя и ветра, одновременно сохраняя воздухопроницаемость. В отличие от других материалов, которые теряют свою эффективность при использовании в неблагоприятных условиях эксплуатации или даже просто после стирки, ткани c мембраной GORE-TEX® невероятно прочны и долговечны.
Материалы GORE-TEX®
Компания W.L. Gore&Associates разработала несколько материалов GORE-TEX®, обладающих определенными свойствами в зависимости от того или иного назначения.
GORE-TEX® Performance Shell обеспечивает полную водонепроницаемость, высокие «дышащие» свойства и комфорт. Идеально подходит для одежды, предназначенной для самого широкого круга outdoor активности.
2-хслойный материал GORE-TEX® Performance Shell состоит из высококачественной наружной ткани, прочно соединенной с мембраной GORE-TEX®, и отдельной подкладки для защиты мембраны.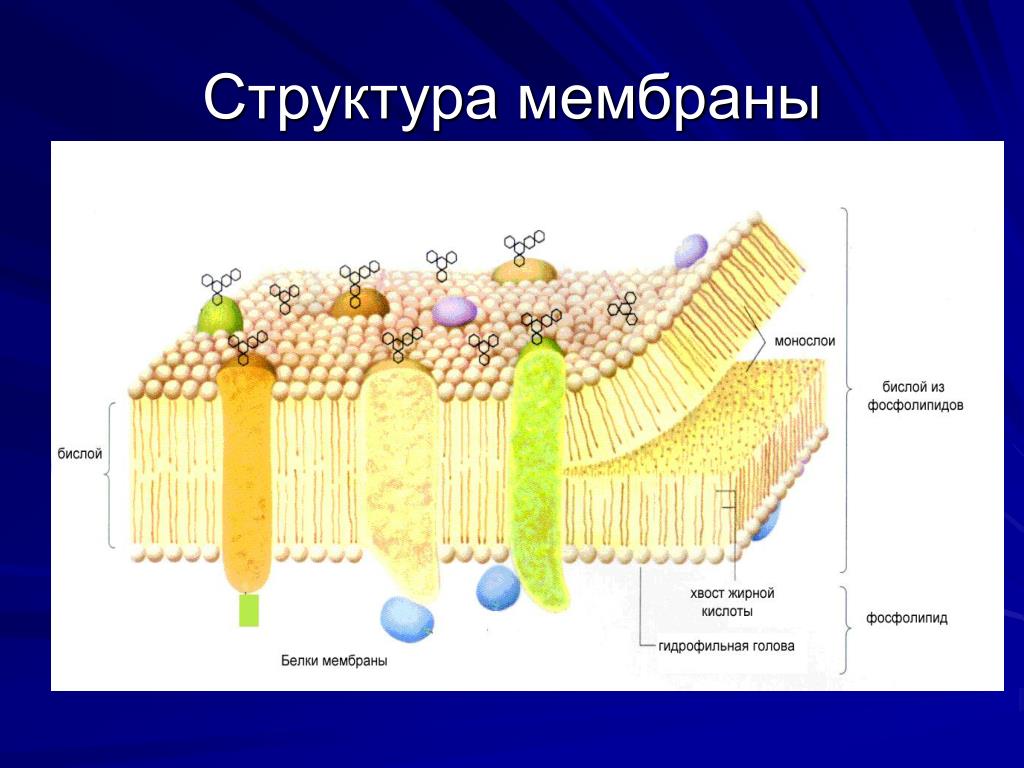
3-хслойный материал GORE-TEX® Performance Shell состоит из высококачественной наружной ткани, прочно соединенной с мембраной GORE-TEX®, и подкладки, прикрепленной к мембране с другой стороны.
GORE-TEX® Pro Shell представляет собой сочетание мембраны с исключительно прочными на разрыв и износ тканями, обладающими высокой «дышащей» способностью. GORE-TEX® Pro Shell специально разработан для обеспечения защиты от непогоды в самых экстремальных условиях.
3-хслойный GORE-TEX® Pro Shell
— Продукт на подведет Вас даже в самых тяжелых погодных условиях
— Одежда очень прочная, прекрасно дышит, а так же защищает от воды и ветра.
— Специальная мембрана GORE-TEX® прочно прикреплена к износостойкому внешнему материалу и подкладке. Это обеспечивает прочность и долгий срок службы одежды даже в экстремальных условиях.
2-хслойный GORE-TEX® Pro Shell
Pro Shell производится из очень износостойкого, прекрасно дышащего материала, который обеспечивает продолжительную защиту от воды и ветра.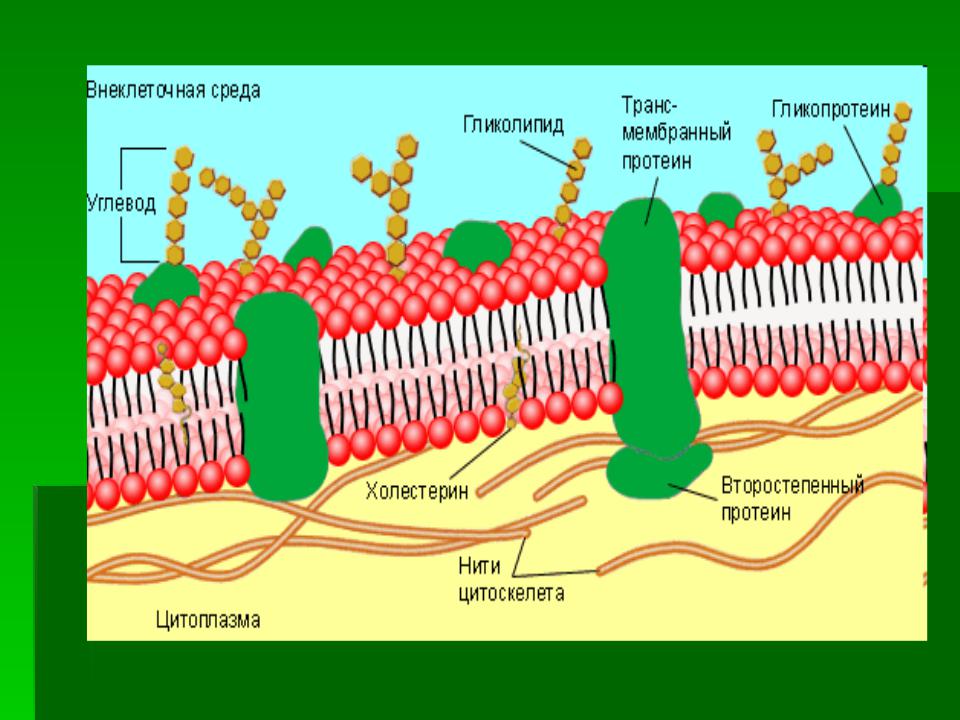 В этом варианте специальная мембрана GORE-TEX® склеивается в ламинат к внутренней стороне внешнего материала. Внутренняя сторона мембраны защищается дополнительной подкладкой. Это гарантирует комфортабельность одежды и ее многосторонность.
В этом варианте специальная мембрана GORE-TEX® склеивается в ламинат к внутренней стороне внешнего материала. Внутренняя сторона мембраны защищается дополнительной подкладкой. Это гарантирует комфортабельность одежды и ее многосторонность.
GORE-TEX® Paclite® Shell сочетает в себе исключительные «дышащие» свойства и ветро-, водонепроницаемость с минимальным весом и компактностью. Идеально подходит для одежды, предназначенной для занятий спортом, где вес и объем экипировки имеет особо важное значение. GORE-TEX® Paclite® Shell имеет специальный защитный слой из маслоотталкивающего вещества и углерода на мембране. Технология нанесения маслоотталкивающего вещества и интеграции углерода ведет к повышению износостойкости мембраны, благодаря чему можно отказаться от дополнительного подкладочного материала. Это позволяет выпускать легкую, удобную и функциональную одежду.
GORE-TEX® Soft Shell представляет собой сочетание мембраны с мягкими тканями, обладающими высокими теплоизолирующими свойствами.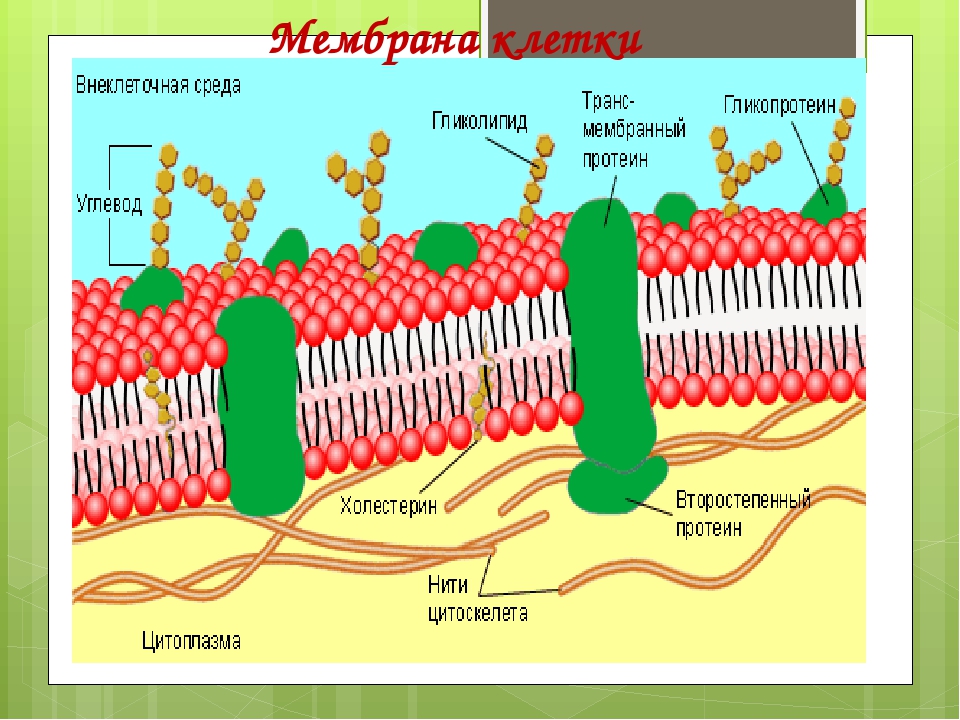 Мембрана располагается между мягким внешним материалом и мягкой подкладкой из флиса. Это сокращает количество необходимых слоев одежды для теплоизоляции и обеспечивает полную свободу движений. Одежда из GORE-TEX® Soft Shell идеально подходит для холодных и влажных погодных условий.
Мембрана располагается между мягким внешним материалом и мягкой подкладкой из флиса. Это сокращает количество необходимых слоев одежды для теплоизоляции и обеспечивает полную свободу движений. Одежда из GORE-TEX® Soft Shell идеально подходит для холодных и влажных погодных условий.
Водонепроницаемая конструкция одежды и обработка швов
Все изделия с использованием мембранных материалов GORE-TEX® создаются только лицензированными производителями. Соблюдение всех технологических требований работы с материалами GORE-TEX®, важнейшим из которых является обработка швов специальной непромокаемой лентой, контролируется непосредственно компанией W.L. Gore&Associates.
Согласно программе контроля качества W.L. Gore&Associates, вся одежда GORE-TEX® конструируется и изготавливается в соответствии со строгими стандартами, гарантирующими защиту от любых погодных условий. Одежда из материалов GORE-TEX® должна иметь специальную конструкцию, предотвращающую проникновение воды через швы и тщательно продуманный дизайн. Компания W.L. Gore&Associates является единственным изготовителем материалов, который использует собственную программу обработки швов.
Компания W.L. Gore&Associates является единственным изготовителем материалов, который использует собственную программу обработки швов.
Тестирование продукции
Для достижения высоких стандартов материалы и готовая одежда GORE-TEX® постоянно проверяются в рамках программы гарантии качества, как в независимых институтах, так и в многочисленных лабораториях и полевых испытаниях.
THE RAIN CHAMBER TEST
Каждая новая модель одежды GORE-TEX® перед запуском в производство должна пройти ряд тестов в дождевой камере Gore (THE RAIN CHAMBER TEST). Дождевая камера Gore — это современная испытательная установка, предназначенная для моделирования различных видов дождя. Благодаря специальным соплам, особым образом размещенным в камере, можно протестировать водонепроницаемость одежды под дождем — от легкой мороси до сильного ливня.
EXTREME WET WEATHER TESTED
Изделия из материалов GORE-TEX®, предназначенные для использования в экстремальных условиях, проходят еще более сложные испытания в дождевой камере, где создаются вертикальные и горизонтальные дождевые потоки. Одежда, прошедшая такое тестирование отвечает наивысшим требованиям защиты в неблагоприятных погодных условиях.
Одежда, прошедшая такое тестирование отвечает наивысшим требованиям защиты в неблагоприятных погодных условиях.
При уходе за изделиями из материалов GORE-TEX®, строго следуйте инструкциям изготовителя на этикетке.
СЕО оптимизация, разработка и реклама сайта в российских и зарубежных поисковиках.
Gore-Tex Одежда Мембранная одежда
Мембранные липиды: где они находятся и как ведут себя
Sud, M. et al. LMSD: база данных структуры LIPID MAPS. Nucleic Acids Res. 35 , D527 – D532 (2007).
CAS PubMed Google ученый
Фейгенсон, Г. У. Фазовое поведение липидных смесей. Nature Chem. Биол. 2 , 560–563 (2006).
CAS Google ученый
Фейгенсон, Г.W. Фазовые границы и биологические мембраны. Annu. Rev. Biophys. Biomol. Struct. 36 , 63–77 (2007).
CAS PubMed PubMed Central Google ученый
Марш, М. и Хелениус, А. Вступление вируса: открытый кунжут. Cell 124 , 729–740 (2006). Исследования клеточной биологии, визуализация живых клеток и системная биология показывают, что многие из множественных и тонко различных путей, которые вирусы животных используют для проникновения в клетки-хозяева, требуют определенных липидов.
CAS PubMed Google ученый
ван Меер, Г. Клеточная липидомика. EMBO J. 24 , 3159–3165 (2005).
CAS PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 6
org/ScholarlyArticle»> 6Марш Д. Профиль бокового давления, нарушение спонтанного искривления, а также включение и конформация белков в мембранах. Biophys.J. 93 , 3884–3899 (2007).
CAS PubMed PubMed Central Google ученый
Dowhan, W. & Bogdanov, M. in Biochemistry of Lipids, Lipoproteins and Membranes Vol. 36 (ред. Вэнс, Д. Э. и Вэнс, Дж. Э.) 1–35 (Elsevier, Амстердам, 2002).
Google ученый
van Meer, G. & Lisman, Q. Транспорт сфинголипидов: плоты и транслокаторы. J. Biol. Chem. 277 , 25855–25858 (2002).
CAS PubMed Google ученый
Хуанг, Дж. И Фейгенсон, Г. В. Микроскопическая модель взаимодействия максимальной растворимости холестерина в липидных бислоях. Biophys. J. 76 , 2142–2157 (1999). Взаимодействие холестерина с мембранными липидами приводит к резким скачкам химического потенциала холестерина из-за гидрофобного взаимодействия, которое заставляет головные группы фосфолипидов защищать холестерин от воды, как описано здесь с помощью зонтичной модели.
И Фейгенсон, Г. В. Микроскопическая модель взаимодействия максимальной растворимости холестерина в липидных бислоях. Biophys. J. 76 , 2142–2157 (1999). Взаимодействие холестерина с мембранными липидами приводит к резким скачкам химического потенциала холестерина из-за гидрофобного взаимодействия, которое заставляет головные группы фосфолипидов защищать холестерин от воды, как описано здесь с помощью зонтичной модели.
CAS PubMed PubMed Central Google ученый
Ali, MR, Cheng, KH & Huang, J. Керамид переводит холестерин из фазы упорядоченного бислоя липидов в кристаллическую фазу в 1-пальмитоил-2-олеоил- sn -глицеро-3-фосфохолине / тройные смеси холестерин / церамид. Биохимия 45 , 12629–12638 (2006).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 11
org/ScholarlyArticle»> 11Мейер цу Херингдорф, Д.И Якобс, К. Х. Лизофосфолипидные рецепторы: передача сигналов, фармакология и регуляция метаболизмом лизофосфолипидов. Биохим. Биофиз. Acta 1768 , 923–940 (2007).
CAS PubMed Google ученый
Фернандис, А.З. и Венк, М.Р. Мембранные липиды как сигнальные молекулы. Curr. Мнение. Липидол. 18 , 121–128 (2007).
CAS PubMed Google ученый
Колесник Р.& Hannun, Ю. А. Керамид и апоптоз. Trends Biochem. Sci. 24 , 224–225 (1999).
CAS PubMed Google ученый
Теппер, А.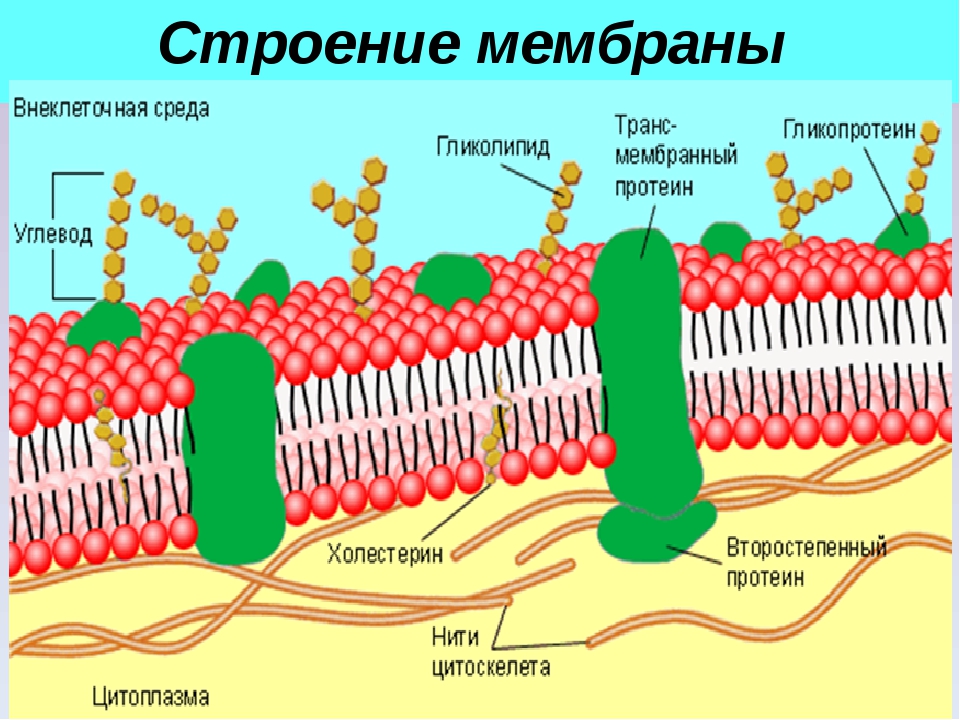 Д. и др. Гидролиз сфингомиелина до церамида во время фазы апоптоза является результатом скремблирования фосфолипидов и изменяет морфологию клеточной поверхности. J. Cell Biol. 150 , 155–164 (2000).
Д. и др. Гидролиз сфингомиелина до церамида во время фазы апоптоза является результатом скремблирования фосфолипидов и изменяет морфологию клеточной поверхности. J. Cell Biol. 150 , 155–164 (2000).
CAS PubMed PubMed Central Google ученый
Megha, Sawatzki, P., Колтер, Т., Биттман, Р. и Лондон, Э. Влияние ацильной цепи N церамида и полярной структуры головной группы на свойства упорядоченных липидных доменов (липидных рафтов). Биохим. Биофиз. Acta 1768 , 2205–2212 (2007).
CAS PubMed PubMed Central Google ученый
Белл Р. М., Баллас Л. М. и Колман Р. А. Топогенез липидов. J. Lipid Res. 22 , 391–403 (1981).
CAS PubMed Google ученый
 org/ScholarlyArticle»> 17
org/ScholarlyArticle»> 17Sprong, H. et al. UDP-галактоза: церамид-галактозилтрансфераза — это интегральный мембранный белок класса I эндоплазматического ретикулума. J. Biol. Chem. 273 , 25880–25888 (1998).
CAS PubMed Google ученый
Русинол, А. Э., Цуй, З., Чен, М. Х. и Вэнс, Дж. Э. Уникальная фракция мембран, ассоциированная с митохондриями, из печени крысы обладает высокой способностью к синтезу липидов и содержит секреторные белки пре-Гольджи, включая возникающие липопротеины. J. Biol. Chem. 269 , 27494–27502 (1994). Предоставляет доказательства стабильных физических ассоциаций между ER и митохондриями, а также определяет биохимические свойства этого компартмента, которые отличаются от отдельных органелл.
CAS PubMed Google ученый
Pichler, H. et al. Субфракция эндоплазматического ретикулума дрожжей ассоциируется с плазматической мембраной и обладает высокой способностью синтезировать липиды. евро. J. Biochem. 268 , 2351–2361 (2001).
CAS PubMed Google ученый
Футерман, А. Х. и Ризман, Х. Все входы и выходы синтеза сфинголипидов. Trends Cell Biol. 15 , 312–318 (2005).
CAS PubMed Google ученый
Хеннебери, А. Л., Райт, М. М. и Макмастер, К. Р.Основные сайты клеточного синтеза фосфолипидов и молекулярные детерминанты жирных кислот и специфичности головной группы липидов. Мол. Биол. Ячейка 13 , 3148–3161 (2002).
CAS PubMed PubMed Central Google ученый
Фолькер Д. Р. Устранение разрывов в транспорте фосфолипидов. Trends Biochem. Sci. 30 , 396–404 (2005). Обобщает биохимические и генетические элементы невезикулярного транспорта фосфолипидов с акцентом на процессы транспорта PtdSer в дрожжах.
CAS PubMed Google ученый
Ди Паоло, Дж. И Де Камилли, П. Фосфоинозитиды в регуляции клеток и мембранной динамике. Природа 443 , 651–657 (2006).
CAS PubMed Google ученый
Geta Tafesse, F. et al. Обе сфингомиелинсинтазы SMS1 и SMS2 необходимы для гомеостаза сфингомиелина и роста клеток HeLa человека. J. Biol. Chem. 282 , 17537–17547 (2007).
Google ученый
Li, Z. et al. Ингибирование сфингомиелинсинтазы (SMS) влияет на накопление внутриклеточного сфингомиелина и липидную организацию плазматической мембраны. Биохим. Биофиз. Acta 1771 , 1186–1194 (2007).
CAS PubMed PubMed Central Google ученый
Кобаяши, Т.и другие. Разделение и характеристика поздних эндосомальных мембранных доменов. J. Biol. Chem. 277 , 32157–32164 (2002).
CAS PubMed Google ученый
Мацуо, Х. и др. Роль LBPA и Alix в формировании мультивезикулярных липосом и организации эндосом. Наука 303 , 531–534 (2004).
CAS PubMed Google ученый
Колтер, Т.И Сандхофф, К. Принципы переваривания лизосомальной мембраны: стимуляция деградации сфинголипидов белками-активаторами сфинголипидов и анионными лизосомальными липидами. Annu. Rev. Cell Dev. Биол. 21 , 81–103 (2005).
CAS PubMed Google ученый
Вэнс Д. и Вэнс Дж. Э. Биохимия липидов, липопротеинов и мембран (Elsevier, Амстердам, 2002).
Google ученый
Нэгл, К.A. et al. Сверхэкспрессия в печени глицерин- sn -3-фосфатацилтрансферазы 1 у крыс вызывает инсулинорезистентность. J. Biol. Chem. 282 , 14807–14815 (2007).
CAS PubMed PubMed Central Google ученый
Choi, J. Y., Wu, W. I. & Voelker, D. R. Фосфатидилсерин декарбоксилазы как генетические и биохимические инструменты для изучения движения фосфолипидов. Анал. Biochem. 347 , 165–175 (2005).
CAS PubMed Google ученый
Даум Г. Липиды митохондрий. Биохим. Биофиз. Acta 822 , 1–42 (1985).
CAS PubMed Google ученый
Штраус, Дж. Ф., Кишида, Т., Кристенсон, Л. К., Фуджимото, Т. и Хирои, белки домена START и внутриклеточный транспорт холестерина в стероидогенных клетках. Мол. Клетка. Эндокринол. 202 , 59–65 (2003).
CAS PubMed Google ученый
Дево П. Ф. и Моррис Р. Трансмембранная асимметрия и латеральные домены в биологических мембранах. Трафик 5 , 241–246 (2004).
CAS PubMed Google ученый
Daleke, D. L. Флиппазы фосфолипидов. Дж.Биол. Chem. 282 , 821–825 (2007).
CAS PubMed Google ученый
Поморски Т. и Менон А. К. Липидные флиппазы и их биологические функции. Ячейка. Мол. Life Sci. 63 , 2908–2921 (2006).
CAS PubMed Google ученый
Энглин, Т. К., Лю, Дж. И Конбой, Дж. С. Легкий липидный триггер в фосфолипидном бислое, индуцированный грамицидином А, измеренный с помощью колебательной спектроскопии суммарной частоты. Biophys. J. 92 , L01 – L03 (2007).
CAS PubMed Google ученый
Пападопулос, А. и др. Активность флиппазы обнаруживается с немечеными липидами по изменению формы гигантских однослойных везикул. J. Biol. Chem. 282 , 15559–15568 (2007).
CAS PubMed Google ученый
López-Montero, I. et al.Быстрое трансбислойное движение церамидов в фосфолипидных везикулах и в эритроцитах человека. J. Biol. Chem. 280 , 25811–25819 (2005).
PubMed Google ученый
Ганонг, Б. Р. и Белл, Р. М. Трансмембранное перемещение аналогов фосфатидилглицерина и диацилглицерина сульфгидрила. Биохимия 23 , 4977–4983 (1984).
CAS PubMed Google ученый
Бай, Дж.И Пагано, Р. Э. Измерение спонтанного переноса и трансбислойного движения липидов, меченных BODIPY, в липидных пузырьках. Биохимия 36 , 8840–8848 (1997).
CAS PubMed Google ученый
Steck, T. L., Ye, J. & Lange, Y. Исследование движения холестерина в мембране эритроцитов с помощью циклодекстрина. Biophys. J. 83 , 2118–2125 (2002).
CAS PubMed PubMed Central Google ученый
Каллис, П.R. et al. Влияние градиентов pH на трансбислойный транспорт лекарств, липидов, пептидов и ионов металлов в большие однослойные везикулы. Биохим. Биофиз. Acta 1331 , 187–211 (1997).
CAS PubMed Google ученый
Кол, М. А., де Крун, А. И., Киллиан, Дж. А. и де Круидж, Б. Трансбислойное движение фосфолипидов в биогенных мембранах. Биохимия 43 , 2673–2681 (2004). Обобщает данные и гипотезы, которые поддерживают общую систему неселективного трансбислойного движения липидов в ЭПР эукариот и в цитоплазматических мембранах бактерий.
CAS PubMed Google ученый
Helenius, J. et al. Для перемещения липид-связанных олигосахаридов через мембрану ER необходим белок Rft1. Nature 415 , 447–450 (2002).
CAS PubMed Google ученый
Алаймо, К.и другие. Два различных, но взаимозаменяемых механизма переворота липид-связанных олигосахаридов. EMBO J. 25 , 967–976 (2006).
CAS PubMed PubMed Central Google ученый
Graham, T.R. Флиппазы и перенос белков, опосредованный пузырьками. Trends Cell Biol. 14 , 670–677 (2004). Обобщает важные взаимосвязи между трафиком везикулярных белков и трансбислойным транспортом фосфолипидов с помощью АТФаз P-типа.
CAS PubMed Google ученый
Pomorski, T. et al. Drs2p-связанные АТФазы P-типа Dnf1p и Dnf2p необходимы для транслокации фосфолипидов через плазматическую мембрану дрожжей и играют роль в эндоцитозе. Мол. Биол. Ячейка 14 , 1240–1254 (2003). Важная статья, определяющая участие АТФаз плазматической мембраны P-типа у дрожжей в трансбислойном перемещении аминоглицерофосфолипидов и их взаимодействии с эндоцитарными процессами.
CAS PubMed PubMed Central Google ученый
Рихоф, В. Р. и Фелькер, Д. Р. Поглощение и использование лизофосфатидилэтаноламина Saccharomyces cerevisiae . J. Biol. Chem. 281 , 36588–36596 (2006).
CAS PubMed Google ученый
Riekhof, W. R. et al. Метаболизм лизо-фосфатидилхолина в Saccharomyces cerevisiae .Роль АТФаз P-типа в транспорте и ацилтрансфераза широкой специфичности в ацилировании. J. Biol. Chem. 21 октября 2007 г. (DOI: 10.1074 / jbc.M706718200)
CAS PubMed Google ученый
Натараджан П., Ван, Дж., Хуа, З. и Грэхэм, Т. Р. Активность транслоказы аминофосфолипидов, связанных с Drs2p, в дрожжевых мембранах Гольджи и связь с функцией in vivo . Proc. Natl. Акад. Sci. США 101 , 10614–10619 (2004).
CAS PubMed Google ученый
Alder-Baerens, N., Lisman, Q., Luong, L., Pomorski, T. и Holthuis, J. C. Потеря P4-АТФаз Drs2p и Dnf3p нарушает транспорт аминофосфолипидов и асимметрию в секреторных пузырьках дрожжей после Гольджи. Мол. Биол. Ячейка 17 , 1632–1642 (2006).
CAS PubMed PubMed Central Google ученый
Ван, Х.и другие. Митохондриальный фактор WAH-1 C. elegans способствует экстернализации фосфатидилсерина в апоптозных клетках посредством фосфолипидной скрамблазы SCRM-1. Nature Cell Biol. 9 , 541–549 (2007).
CAS PubMed Google ученый
Züllig, S. et al. Аминофосфолипидная транслоказа ТАТ-1 способствует экспозиции фосфатидилсерина во время апоптоза C. elegans . Curr. Биол. 17 , 994–999 (2007).
PubMed Google ученый
van Meer, G. & Simons, K. Функция плотных контактов в поддержании различий в липидном составе между апикальными и базолатеральными доменами клеточной поверхности клеток MDCK. EMBO J. 5 , 1455–1464 (1986).
CAS PubMed PubMed Central Google ученый
Янг, W. W. Jr, Lutz, M.S. & Blackburn, W. A. Эндогенные гликосфинголипиды перемещаются к поверхности клетки со скоростью, соответствующей оценкам объемного потока. J. Biol. Chem. 267 , 12011–12015 (1992).
CAS PubMed Google ученый
Baumann, N.A. et al. Транспорт вновь синтезированного стерола к обогащенной стерином плазматической мембране происходит посредством невезикулярного уравновешивания. Биохимия 44 , 5816–5826 (2005).
CAS PubMed Google ученый
Саймонс К. и ван Меер Г. Сортировка липидов в эпителиальных клетках. Биохимия 27 , 6197–6202 (1988).
CAS PubMed Google ученый
Halter, D. et al. Пре- и пост-Гольджи-транслокация глюкозилцерамида в синтезе гликосфинголипидов J. Cell Biol. 179 , 101–115 (2007). предполагает, что поздний белок Гольджи FAPP2 транспортирует GlcCer, который предназначен для синтеза сложных гликолипидов, обратно в ER, тогда как перемещение GlcCer на поверхность клетки зависит от протонного градиента.
CAS PubMed PubMed Central Google ученый
Smith, D.C. et al. Связь шига-подобного токсина с устойчивыми к детергентам мембранами модулируется глюкозилцерамидом и является важным требованием эндоплазматического ретикулума для цитотоксического действия. Мол. Биол. Ячейка 17 , 1375–1387 (2006).
CAS PubMed PubMed Central Google ученый
Sharma, D. K. et al. Гликосфинголипиды, интернализованные через кавеолярный эндоцитоз, быстро сливаются с клатриновым путем в ранних эндосомах и образуют микродомены для рециклинга. J. Biol. Chem. 278 , 7564–7572 (2003). Демонстрирует неравномерное распределение флуоресцентных фосфолипидов в отдельных эндосомах.
CAS PubMed Google ученый
Wang, T. Y. & Silvius, J. R. Различные сфинголипиды обнаруживают дифференциальное разделение на сфинголипидные / богатые холестерином домены в липидных бислоях. Biophys. J. 79 , 1478–1489 (2000).
CAS PubMed PubMed Central Google ученый
Сингх Р. Д. и др. Ингибирование захвата кавеолой, инфекции SV40 и передачи сигналов β1-интегрина неприродным стереоизомером гликосфинголипида. J. Cell Biol. 176 , 895–901 (2007).
CAS PubMed PubMed Central Google ученый
Sleight, R.G. & Pagano, R.E. Быстрое появление вновь синтезированного фосфатидилэтаноламина на плазматической мембране. J. Biol. Chem. 258 , 9050–9058 (1983).
CAS PubMed Google ученый
Каплан, М.Р. и Симони, Р. Д. Внутриклеточный транспорт фосфатидилхолина к плазматической мембране. J. Cell Biol. 101 , 441–445 (1985).
CAS PubMed Google ученый
Фолькер Д. Р. Характеристика синтеза и транслокации фосфатидилсерина в проницаемых клетках животных. J. Biol. Chem. 265 , 14340–14346 (1990).
CAS PubMed Google ученый
Вэнс, Дж.E., Aasman, E. J. & Szarka, R. Brefeldin A не подавляет перемещение фосфатидилэтаноламина от участков его синтеза к поверхности клетки. J. Biol. Chem. 266 , 8241–8247 (1991).
CAS PubMed Google ученый
Wu, W. I. & Voelker, D. R. Характеристика транспорта фосфатидилсерина к локусу фосфатидилсерин декарбоксилазы 2 в пермеабилизированных дрожжах. Дж.Биол. Chem. 276 , 7114–7121 (2001).
CAS PubMed Google ученый
Schumacher, M. M., Choi, J. Y., Voelker, D. R. Транспорт фосфатидилсерина в митохондрии регулируется убиквитинированием. J. Biol. Chem. 277 , 51033–51042 (2002).
CAS PubMed Google ученый
Пападопулос, В.и другие. Белок-транслокатор (18 кДа): новая номенклатура бензодиазепинового рецептора периферического типа, основанная на его структуре и молекулярной функции. Trends Pharmacol. Sci. 27 , 402–409 (2006). Обобщает основные белковые компоненты, участвующие в невезикулярном импорте холестерина в митохондрии клеток, участвующих в синтезе стероидных гормонов.
CAS PubMed Google ученый
Ханада, К.и другие. Молекулярный аппарат невезикулярного движения церамида. Природа 426 , 803–809 (2003). Важная статья, определяющая генетические и биохимические механизмы невезикулярного транспорта церамида между ЭПР и аппаратом Гольджи.
CAS Google ученый
Варнок, Д. Э., Лутц, М. С., Блэкберн, В. А., Янг, В. В. Младший, и Баензигер, Дж. У. Транспорт вновь синтезированного глюкозилцерамида к плазматической мембране не по пути Гольджи. Proc. Natl. Акад. Sci. США 91 , 2708–2712 (1994).
CAS PubMed Google ученый
Trotter, P.J., Wu, W. I., Pedretti, J., Yates, R. & Voelker, D. R. Генетический скрининг мутантов, переносящих аминофосфолипиды, идентифицирует фосфатидилинозитол-4-киназу, STT4p, как важный компонент в метаболизме фосфатидилсерина. J. Biol. Chem. 273 , 13189–13196 (1998).
CAS PubMed Google ученый
Wu, W. I. & Voelker, D. R. Восстановление транспорта фосфатидилсерина от химически определенных донорских мембран на фосфатидилсерин декарбоксилазу 2 вовлекает в процесс специфические липидные домены. J. Biol. Chem. 279 , 6635–6642 (2004).
CAS PubMed Google ученый
Райчаудхури, С., Im, Y.J., Hurley, J.H. & Prinz, W.A. Невезикулярное перемещение стеролов от плазматической мембраны к ER требует оксистерин-связывающих белков, связанных с белками, и фосфоинозитидов. J. Cell Biol. 173 , 107–119 (2006).
CAS PubMed PubMed Central Google ученый
D’Angelo, G. et al. Синтез гликосфинголипидов требует передачи глюкозилцерамида FAPP2. Nature 449 , 62–67 (2007). Показывает, что FAPP2, белок, который связан с генерацией транспортных носителей от Гольджи к плазматической мембране, по-видимому, является белком-переносчиком GlcCer, играющим ключевую роль в комплексном синтезе GSL.
CAS Google ученый
Awai, K., Xu, C., Tamot, B. & Benning, C. Связывающий фосфатидную кислоту белок мембраны внутренней оболочки хлоропласта, участвующий в переносе липидов. Proc.Natl Acad. Sci. США 103 , 10817–10822 (2006). Определяет компоненты транспорта и межмембранного распознавания для перемещения фосфолипидов между внешней и внутренней мембранами хлоропластов.
CAS PubMed Google ученый
Тефсен, Б., Геуртсен, Дж., Бекерс, Ф., Томмассен, Дж. И де Кок, Х. Транспорт липополисахаридов к бактериальной внешней мембране в сферопластах. J. Biol. Chem. 280 , 4504–4509 (2005).
CAS PubMed Google ученый
Мусли, К. Дж., Терьяр, К. Р., Винсент-Поуп, П. и Банкайтис, В. А. Sec14-суперсемейство и регуляторный интерфейс между метаболизмом фосфолипидов и мембранным переносом. Биохим. Биофиз. Acta 1771 , 727–736 (2007).
CAS PubMed PubMed Central Google ученый
Литвак, В., Dahan, N., Ramachandran, S., Sabanay, H. & Lev, S. Поддержание уровня диацилглицерина в аппарате Гольджи с помощью белка Nir2 имеет решающее значение для секреторной функции Гольджи. Nature Cell Biol. 7 , 225–234 (2005).
CAS PubMed Google ученый
Черномордик, Л., Козлов, М. М., Циммерберг, Дж. Липиды в слиянии биологических мембран. J. Membr. Биол. 146 , 1–14 (1995).
CAS PubMed Google ученый
Шемеш, Т., Луини, А., Малхотра, В., Бургер, К. Н. и Козлов, М. М. Сужение трубчатых носителей Гольджи перед делением, вызванное локальным метаболизмом липидов: теоретическая модель. Biophys. J. 85 , 3813–3827 (2003).
CAS PubMed PubMed Central Google ученый
Геннис Р.Б. Биомембраны. Молекулярная структура и функция (Springer Verlag, New York, 1989).
Google ученый
Takamori, S. et al. Молекулярная анатомия органеллы трафика. Cell 127 , 831–846 (2006). Тщательная реконструкция синаптических пузырьков показывает, что холестерин и фосфолипиды (молярное отношение 0,8) покрывают ~ 70% поверхности, в то время как трансмембранные спирали покрывают 20% с соотношением липид / белок 0.75 (по массе).
CAS Google ученый
Dietrich, C., Volovyk, ZN, Levi, M., Thompson, NL & Jacobson, K. Разделение Thy-1, GM1 и сшитых аналогов фосфолипидов в липидные рафты, восстановленные в монослоях поддерживаемой модели мембраны . Proc. Natl Acad. Sci. США 98 , 10642–10647 (2001).
CAS PubMed Google ученый
Морс, С.A. Базальты и фазовые диаграммы (Springer-Verlag, New York, 1980).
Google ученый
Партон Р.Г. Ультраструктурная локализация ганглиозидов: GM1 сосредоточен в кавеолах. J. Histochem. Cytochem. 42 , 155–166 (1994).
CAS PubMed Google ученый
Kusumi, A. et al. Сдвиг парадигмы концепции плазматической мембраны от двумерной континуальной жидкости к разделенной жидкости: высокоскоростное отслеживание одной молекулы мембранных молекул. Annu. Rev. Biophys. Biomol. Struct. 34 , 351–378 (2005). Липиды и белки биологических мембран разделены на участки размером в несколько десятков нанометров, которые имеют различные молекулярные компоненты и свойства.
CAS PubMed Google ученый
Лагерхольм, Б. К., Вайнреб, Г. Э., Якобсон, К. и Томпсон, Н. Л. Обнаружение микродоменов в интактных клеточных мембранах. Annu.Rev. Phys. Chem. 56 , 309–336 (2005).
CAS PubMed Google ученый
Фейгенсон, Г. У. и Бубольц, Дж. Т. Тройная фазовая диаграмма дипальмитоил-ПК / дилауроил-ПК / холестерин: формирование наноскопических доменов под действием холестерина. Biophys. J. 80 , 2775–2788 (2001).
CAS PubMed PubMed Central Google ученый
Медер, Д., Морено, М. Дж., Веркаде, П., Ваз, В. Л. и Саймонс, К. Фазовое сосуществование и взаимосвязь в апикальной мембране поляризованных эпителиальных клеток. Proc. Natl Acad. Sci. США 103 , 329–334 (2006).
CAS PubMed Google ученый
Кусуми, А., Кояма-Хонда, И. и Сузуки, К. Молекулярная динамика и взаимодействия для создания индуцированных стимуляцией стабилизированных плотов из небольших нестабильных стационарных плотов. Трафик 5 , 213–230 (2004).
CAS PubMed Google ученый
Боллинджер, К. Р., Тайхграбер, В. и Гулбинс, Е. Обогащенные церамидом мембранные домены. Биохим. Биофиз. Acta 1746 , 284–294 (2005).
CAS PubMed Google ученый
Roux, A. et al. Роль кривизны и фазового перехода в сортировке липидов и делении мембранных канальцев. EMBO J. 24 , 1537–1545 (2005).
CAS PubMed PubMed Central Google ученый
Chiantia, S., Kahya, N., Ries, J. & Schwille, P. Эффекты церамида на жидкоупорядоченные домены исследуются с помощью одновременного АСМ и FCS. Biophys. J. 90 , 4500–4508 (2006).
CAS PubMed PubMed Central Google ученый
Сот, Дж., Багатолли, Л. А., Гони, Ф. М. и Алонсо, А. Устойчивые к детергентам, обогащенные церамидами домены в бислоях сфингомиелина / церамида. Biophys. J. 90 , 903–914 (2006).
CAS PubMed Google ученый
Анишкин А., Сухарев С. и Коломбини М. Поиск молекулярной структуры трансмембранных церамидных каналов. Biophys. J. 90 , 2414–2426 (2006).
CAS PubMed PubMed Central Google ученый
Дитрих, К., Янг, Б., Фудзивара, Т., Кусуми, А. и Якобсон, К. Связь липидных рафтов с переходными зонами удержания, обнаруженными с помощью отслеживания одиночных частиц. Biophys. J. 82 , 274–284 (2002).
CAS PubMed PubMed Central Google ученый
Сенгупта, П., Бэрд, Б. и Холовка, Д. Липидные рафты, разделение фаз жидкость / жидкость и их значение для структуры и функции плазматической мембраны. Семин.Cell Dev. Биол. 18 , 583–590 (2007).
CAS PubMed PubMed Central Google ученый
Диббл, А. Р. и Фейгенсон, Г. В. Обнаружение сосуществующих жидких фосфолипидных фаз посредством равновесного связывания Ca 2+ : бедное пептидом Lα и богатое пептидом HII сосуществование фаз в дисперсиях грамицидина A ‘/ фосфолипидов. Биохимия 33 , 12945–12953 (1994).
CAS PubMed Google ученый
Льюис, Р.N. et al. Исследования минимальной гидрофобности α-спиральных пептидов, необходимых для поддержания стабильной трансмембранной ассоциации с фосфолипидными двухслойными мембранами. Биохимия 46 , 1042–1054 (2007).
CAS PubMed PubMed Central Google ученый
Якобсон, К., Моуритсен, О. Г. и Андерсон, Р. Г. Липидные плоты: на перекрестке между клеточной биологией и физикой. Nature Cell Biol. 9 , 7–14 (2007). Предлагает модель, согласно которой трансмембранные спирали покрывают 15% поверхности и физически контактируют с 30% мембранных липидов, называемых липидами оболочки. Получив сигнал, белки контролируют фазовое поведение, комбинируя свою оболочку с аналогичными липидными оболочками других белков.
CAS PubMed Google ученый
Shogomori, H. et al. Пальмитоилирование и взаимодействия внутриклеточных доменов способствуют нацеливанию линкера на рафт для активации Т-клеток. J. Biol. Chem. 280 , 18931–18942 (2005).
CAS PubMed Google ученый
Браун Д. А. Липидные рафты, устойчивые к детергентам мембраны и сигналы нацеливания на рафты. Физиология 21 , 430–439 (2006). Использование устойчивости к детергентам для изучения свойств биологических мембран дает косвенную информацию о предпочтениях белков и липидных фаз, но не дает моментального снимка фактического фазового поведения.
CAS PubMed Google ученый
Эпанд, Р. М. Холестерин и взаимодействие белков с мембранными доменами. Прог. Lipid Res. 45 , 279–294 (2006).
CAS PubMed Google ученый
Хэнкок, Дж. Ф. Липидные плоты: спорны только с упрощенной точки зрения. Nature Rev. Mol. Cell Biol. 7 , 456–462 (2006). Критическое обсуждение липидных рафтов, подчеркивающее, что белок-белковые взаимодействия вносят основной вклад в стабильность липидных доменов, и что белки и специфические липиды, такие как холестерин, могут накапливаться на границах доменов и влиять на них.
CAS Google ученый
Лондон, Э. и Фейгенсон, Г. В. Тушение флуоресценции в модельных мембранах. 2. Определение местного липидного окружения аденозинтрифосфатазы кальция из саркоплазматического ретикулума. Биохимия 20 , 1939–1948 (1981).
CAS PubMed Google ученый
Кэффри М. и Фейгенсон Дж. У. Тушение флуоресценции в модельных мембранах. 3. Взаимосвязь между активностью фермента аденозинтрифосфатазы кальция и сродством белка к фосфатидилхолинам с различными характеристиками ацильной цепи. Биохимия 20 , 1949–1961 (1981).
CAS PubMed PubMed Central Google ученый
Эсманн, М.И Марш Д. Липидно-белковые взаимодействия с Na, K-АТФазой. Chem. Phys. Липиды 141 , 94–104 (2006).
CAS PubMed Google ученый
Субиас, О., Тиг, У. Э. и Гавриш, К. Доказательства специфичности липид-родопсиновых взаимодействий. J. Biol. Chem. 281 , 33233–33241 (2006).
CAS PubMed Google ученый
Андерсен, О.С. и Кёпп, Р. Э. 2-й. Толщина бислоя и функция мембранного белка: энергетическая перспектива. Annu. Rev. Biophys. Biomol. Struct. 36 , 107–130 (2007).
CAS PubMed Google ученый
Sharma, P. et al. Наноразмерная организация множественных GPI-заякоренных белков в мембранах живых клеток. Cell 116 , 577–589 (2004).
CAS PubMed Google ученый
Ректенвальд, Д.Дж. И МакКоннелл, Х. М. Фазовые равновесия в бинарных смесях фосфатидилхолина и холестерина. Биохимия 20 , 4505–4510 (1981).
CAS PubMed Google ученый
Wang, T. Y. & Silvius, J. R. Холестерин не индуцирует сегрегацию жидкоупорядоченных доменов в двухслойных слоях, моделирующих внутренний листок плазматической мембраны. Biophys. J. 81 , 2762–2773 (2001).
CAS PubMed PubMed Central Google ученый
Кисслинг, В., Крейн, Дж. М. и Тамм, Л. К. Трансбиллойные эффекты рафтообразных липидных доменов в асимметричных плоских бислоях, измеренные с помощью трекинга одиночных молекул. Biophys. J. 91 , 3313–3326 (2006).
CAS PubMed PubMed Central Google ученый
ван Меер, Г., Halter, D., Sprong, H., Somerharju, P. & Egmond, M.R. Переносчики липидов ABC: экструдеры, флиппазы или активаторы без флопсов? FEBS Lett. 580 , 1171–1177 (2006).
CAS PubMed Google ученый
Витч, С. Л. и Келлер, С. Л. Разделение жидких фаз в гигантских везикулах тройных смесей фосфолипидов и холестерина. Biophys. J. 85 , 3074–3083 (2003).
CAS PubMed PubMed Central Google ученый
Вич, С. Л., Полозов, И. В., Гавриш, К. и Келлер, С. Л. Жидкие домены в везикулах, исследованные методами ЯМР и флуоресцентной микроскопии. Biophys. J. 86 , 2910–2922 (2004).
CAS PubMed PubMed Central Google ученый
Wassall, S. R. et al. Приказ от беспорядка, загоняющий холестерин хаотическими липидами.Роль полиненасыщенных липидов в формировании мембранного рафта. Chem. Phys. Липиды 132 , 79–88 (2004).
CAS PubMed Google ученый
Veatch, S. L., Gawrisch, K. & Keller, S. L. Разрыв в смешиваемости с замкнутым контуром и количественные связующие линии в тройных мембранах, содержащих дифитаноил PC. Biophys. J. 90 , 4428–4436 (2006).
CAS PubMed PubMed Central Google ученый
Бахт, О., Pathak, P. & London, E. Влияние структуры липидов, способствующих образованию неупорядоченных доменов, на стабильность холестерин-содержащих упорядоченных доменов (липидных рафтов): идентификация множественных механизмов стабилизации рафтов. Biophys. J. 93 , 4307–4318 (2007). Полиненасыщенные ацильные цепи мембранных липидов могут эффективно управлять образованием мембранных рафтов из-за особенно плохой упаковки холестерином.
CAS PubMed PubMed Central Google ученый
Куцерка, Н., Тристрам-Нэгл, С. и Нэгл, Дж. Ф. Более пристальный взгляд на структуру полностью гидратированных бислоев ДПФХ жидкой фазы. Biophys. J. 90 , L83 – L85 (2006).
CAS PubMed PubMed Central Google ученый
Лю К., Хуа З., Непуте Дж. А. и Грэхэм Т. Р. Р4-АТФазы дрожжей Drs2p и Dnf1p являются основными грузами эндоцитарного пути NPFXD / Sla1p. Мол. Биол. Cell 18 , 487–500 (2007).
CAS PubMed PubMed Central Google ученый
Структура мембраны | Биология для майоров I
Опишите структуру и функцию мембран, особенно бислоя фосфолипидов.
В результате мы узнаем о структуре мембран.
Цели обучения
- Опишите строение клеточных мембран
- Определить компоненты клеточной мембраны, включая фосфолипиды, холестерин, белки и углеводы
- Объясните, почему гидрофильные вещества не могут проходить через клеточную мембрану
Структура клеточной мембраны
Плазматическая мембрана клетки определяет границу клетки и определяет характер ее контакта с окружающей средой.Клетки исключают одни вещества, поглощают другие и выделяют третьи в контролируемых количествах. Плазматические мембраны ограничивают границы клеток, но они не статичны, а динамичны и постоянно изменяются. Плазматическая мембрана должна быть достаточно гибкой, чтобы определенные клетки, такие как эритроциты и лейкоциты, могли изменять форму при прохождении через узкие капилляры. Это наиболее очевидные функции плазматической мембраны. Кроме того, на поверхности плазматической мембраны есть маркеры, которые позволяют клеткам узнавать друг друга, что жизненно важно, поскольку ткани и органы формируются на раннем этапе развития, и которые позже играют роль в различении «я» и «не-я». иммунный ответ.
Плазматическая мембрана также несет рецепторы, которые являются местами прикрепления определенных веществ, взаимодействующих с клеткой. Каждый рецептор устроен так, чтобы связываться с определенным веществом. Например, поверхностные рецепторы мембраны создают изменения внутри, такие как изменения ферментов метаболических путей. Эти метаболические пути могут иметь жизненно важное значение для обеспечения клетки энергией, выработки определенных веществ для клетки или расщепления клеточных отходов или токсинов для утилизации.Рецепторы на внешней поверхности плазматической мембраны взаимодействуют с гормонами или нейротрансмиттерами и позволяют передавать свои сообщения в клетку. Некоторые сайты распознавания используются вирусами как точки прикрепления. Хотя они очень специфичны, патогены, такие как вирусы, могут развиваться, чтобы использовать рецепторы, чтобы проникнуть в клетку, имитируя конкретное вещество, которое рецептор должен связывать. Эта специфичность помогает объяснить, почему вирус иммунодефицита человека (ВИЧ) или любой из пяти типов вирусов гепатита вторгаются только в определенные клетки.
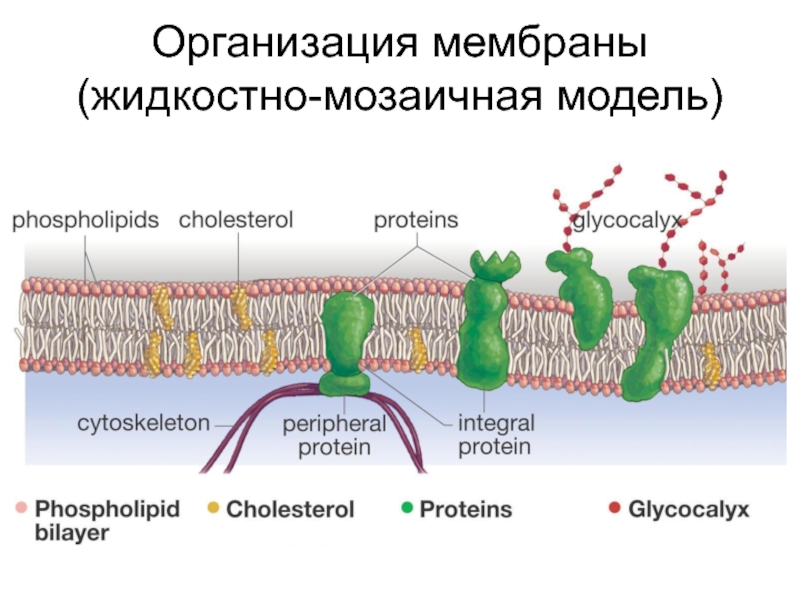
Жидкая мозаика Модель
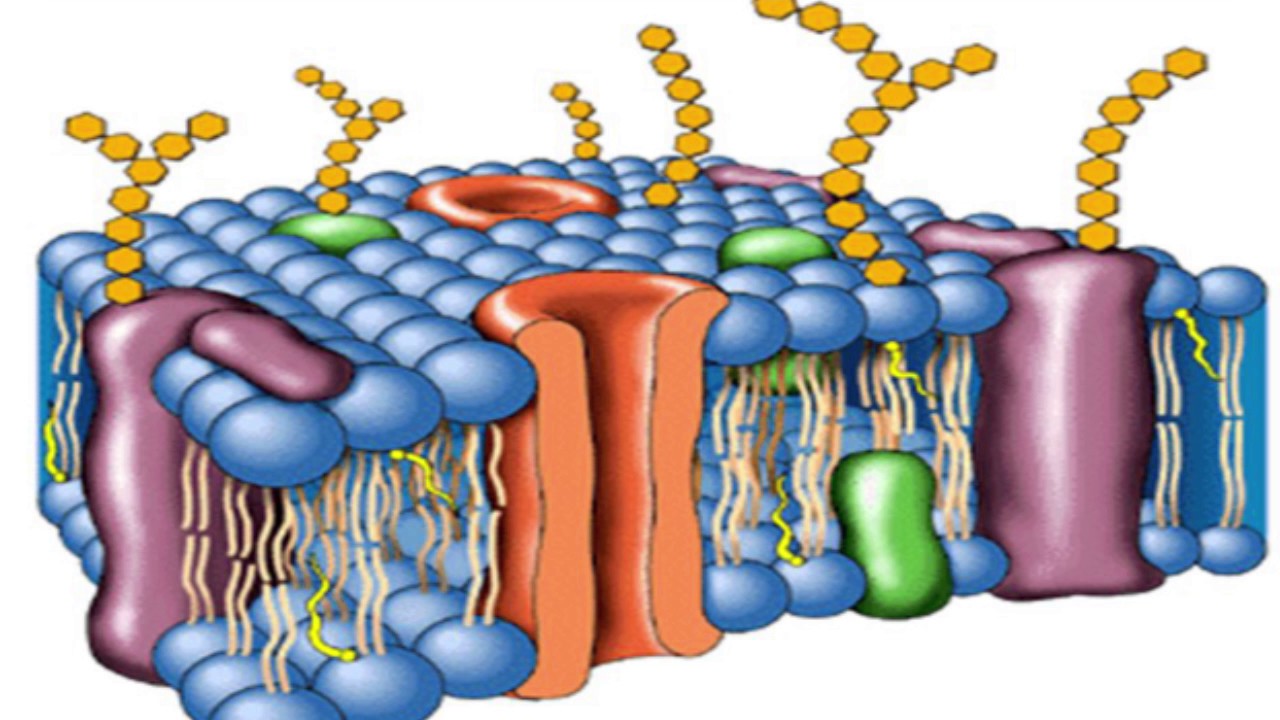
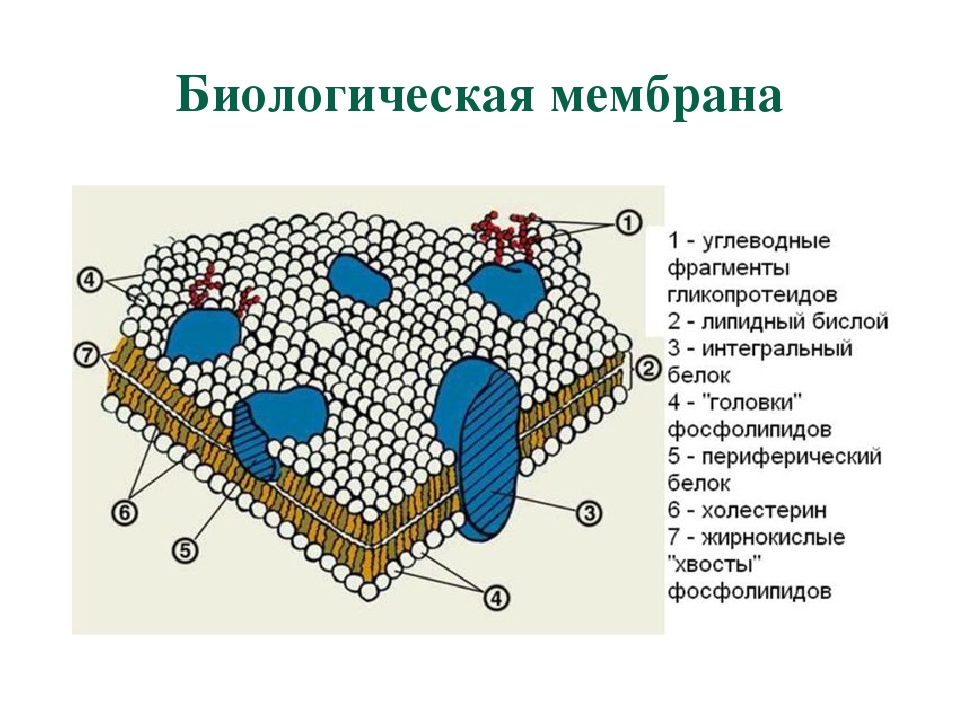
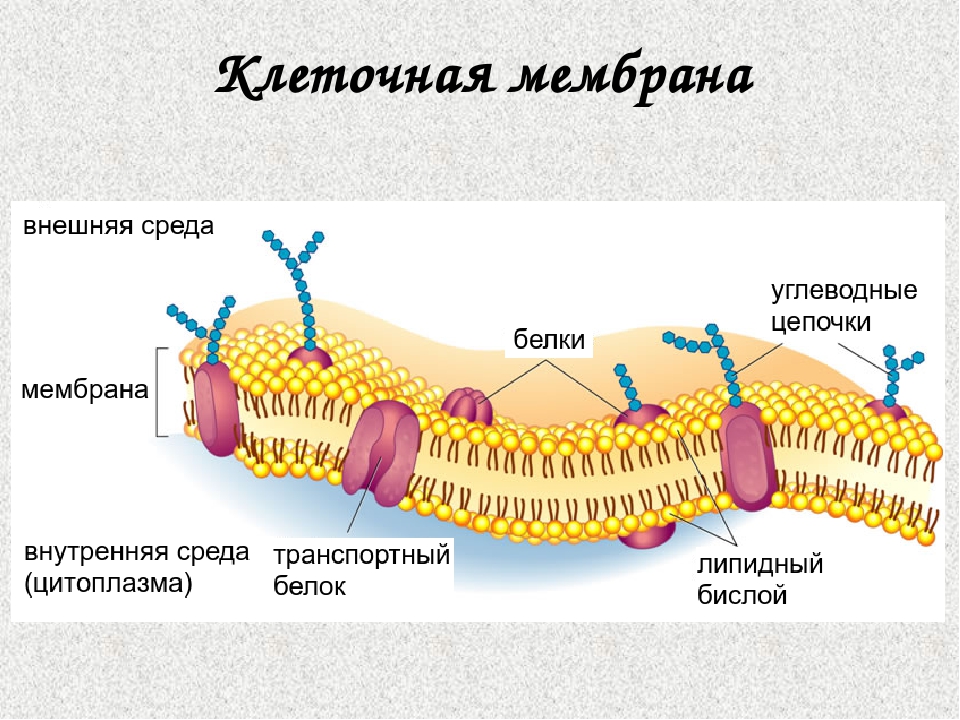
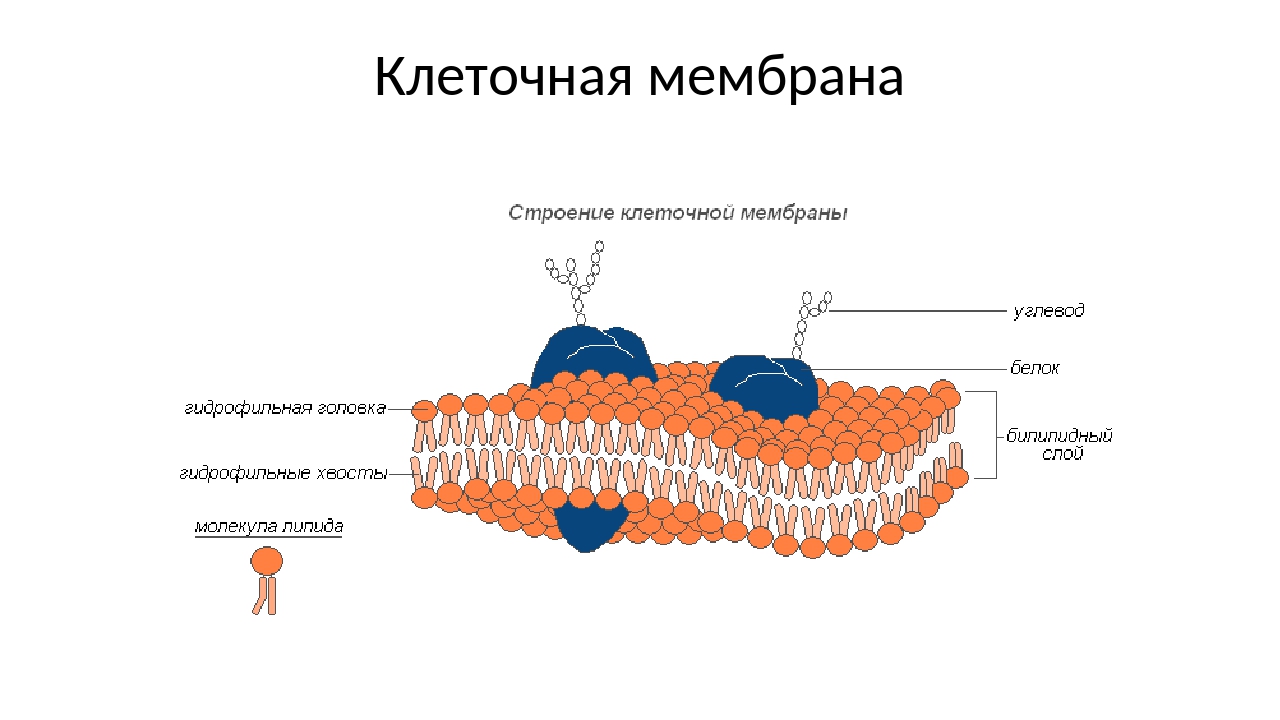
В 1972 году С. Дж. Сингер и Гарт Л. Николсон предложили новую модель плазматической мембраны, которая, по сравнению с более ранним пониманием, лучше объясняла как микроскопические наблюдения, так и функцию плазматической мембраны. Это была модель жидкой мозаики . Модель со временем несколько эволюционировала, но по-прежнему лучше всего объясняет структуру и функции плазматической мембраны в том виде, в каком мы их теперь понимаем. Модель жидкой мозаики описывает структуру плазматической мембраны как мозаику компонентов, включая фосфолипиды, холестерин, белки и углеводы, в которых компоненты могут течь и менять положение, сохраняя при этом базовую целостность мембраны.Как молекулы фосфолипидов, так и встроенные белки способны быстро и латерально диффундировать в мембрану (рис. 1). Текучесть плазматической мембраны необходима для активности определенных ферментов и транспортных молекул внутри мембраны. Плазменные мембраны имеют толщину от 5 до 10 нм. Для сравнения, красные кровяные тельца человека, видимые с помощью световой микроскопии, имеют толщину примерно 8 мкм, что примерно в 1000 раз толще плазматической мембраны.
Рис. 1. Жидкая мозаичная модель структуры плазматической мембраны описывает плазматическую мембрану как жидкую комбинацию фосфолипидов, холестерина, белков и углеводов.
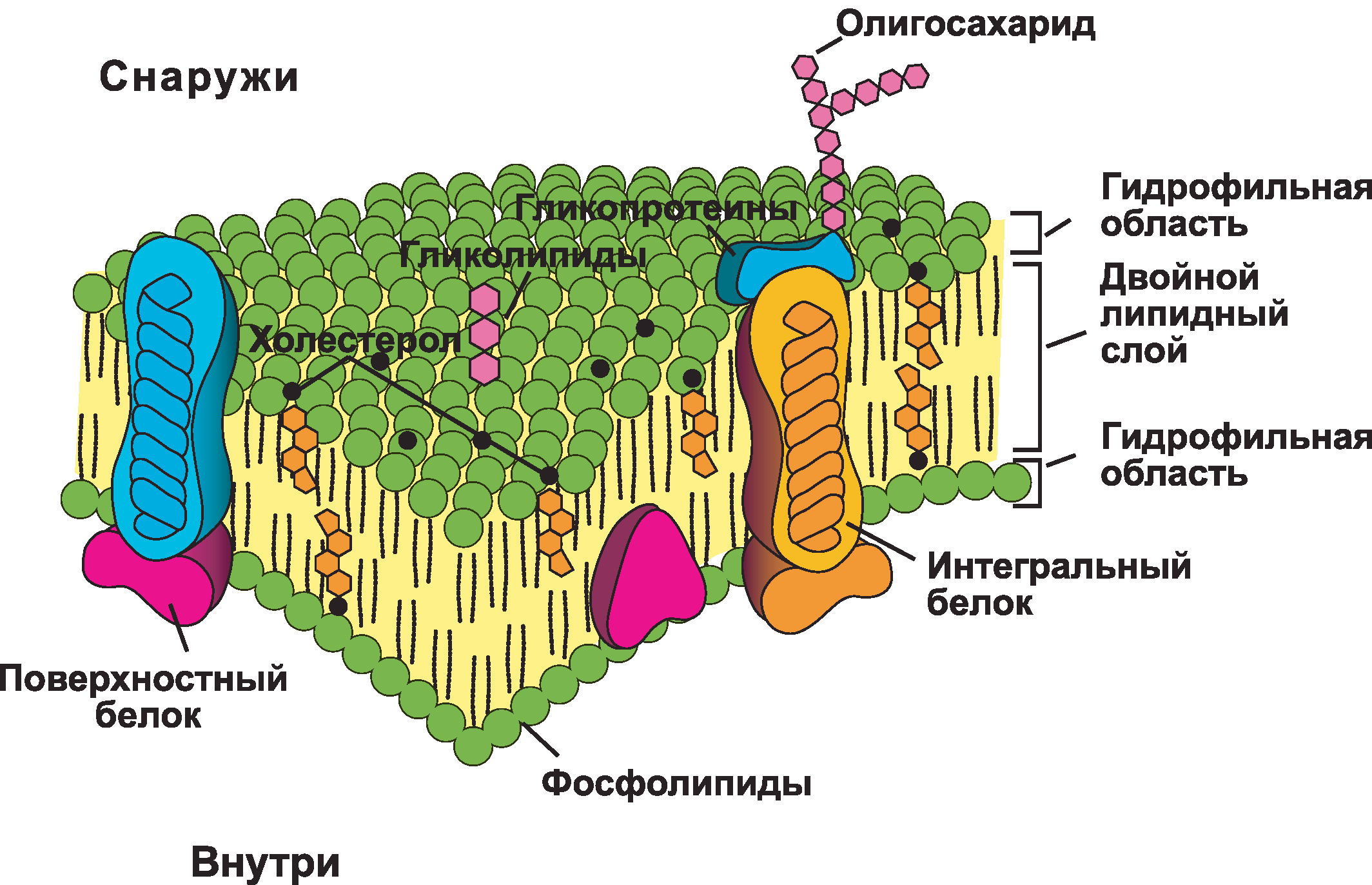
Плазматическая мембрана состоит в основном из бислоя фосфолипидов со встроенными белками, углеводами, гликолипидами и гликопротеинами, а в клетках животных — холестерином. Количество холестерина в плазматических мембранах животных регулирует текучесть мембраны и изменяется в зависимости от температуры окружающей среды клетки. Другими словами, холестерин действует как антифриз на клеточной мембране, и его больше у животных, живущих в холодном климате.
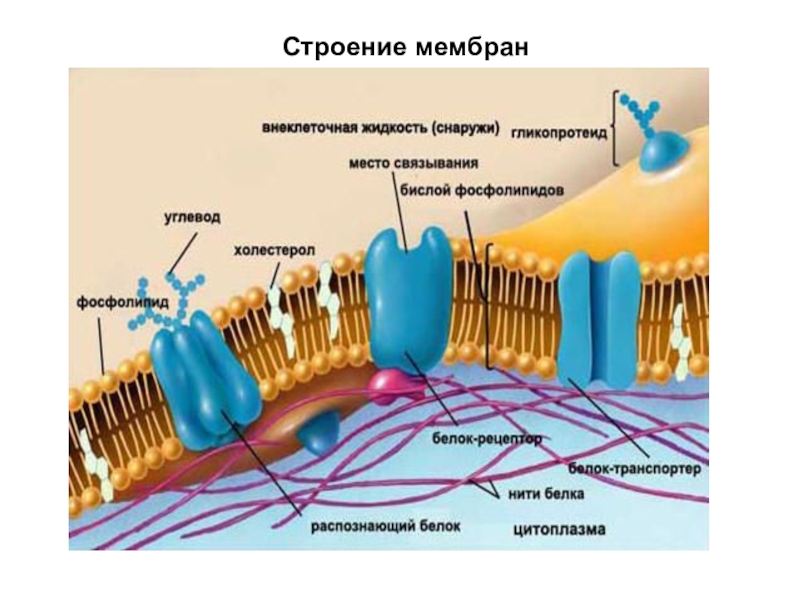
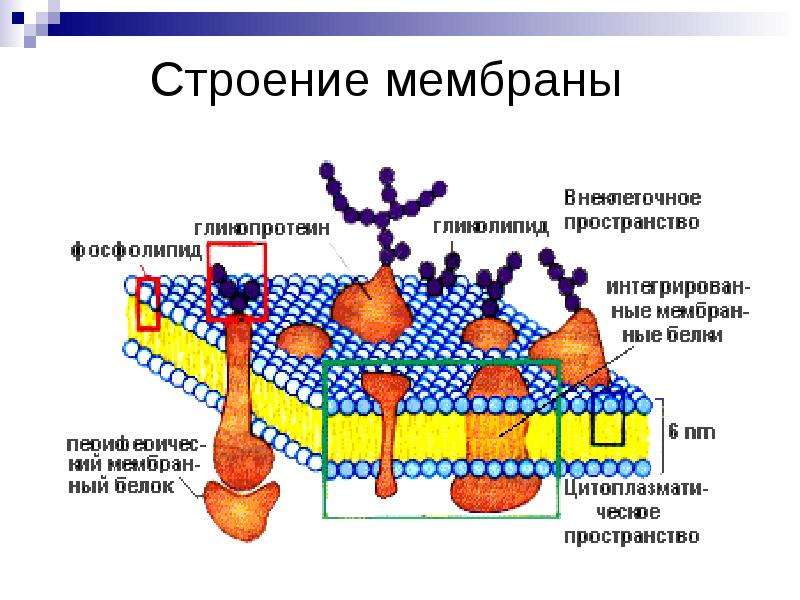
Основная ткань мембраны состоит из двух слоев молекул фосфолипидов, и полярные концы этих молекул (которые выглядят как набор шариков в изображении модели художником) (рис. 1) находятся в контакте с водной жидкостью. внутри и снаружи клетки.Таким образом, обе поверхности плазматической мембраны гидрофильны («водолюбивы»). Напротив, внутренняя часть мембраны между двумя ее поверхностями представляет собой гидрофобную («ненавидящую воду») или неполярную область из-за хвостов жирных кислот. Эта область не имеет притяжения для воды или других полярных молекул (мы обсудим это далее на следующей странице).
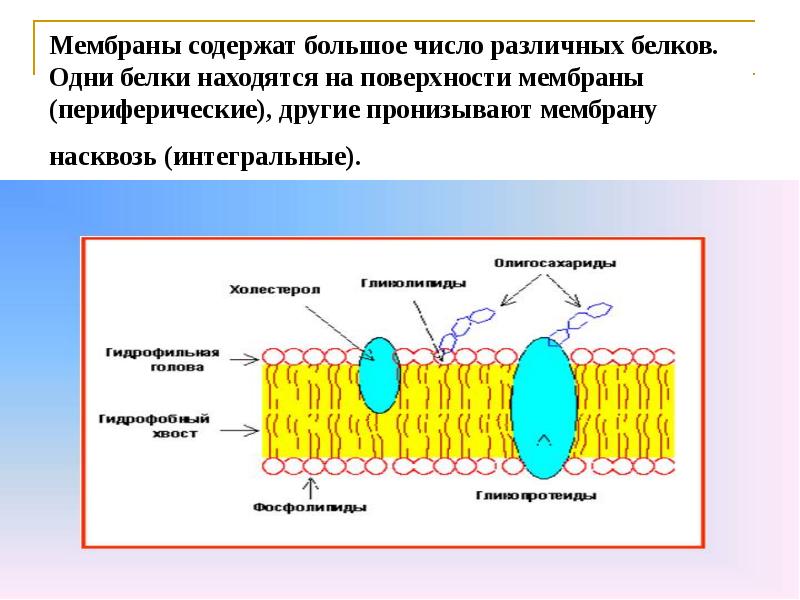
Белки составляют второй по величине химический компонент плазматических мембран. Интегральные белки встроены в плазматическую мембрану и могут охватывать всю или часть мембраны.Интегральные белки могут служить в качестве каналов или насосов для перемещения материалов в клетку или из нее. Периферические белки находятся на внешней или внутренней поверхности мембран, прикрепленные либо к интегральным белкам, либо к молекулам фосфолипидов. Как интегральные, так и периферические белки могут служить ферментами, структурными прикреплениями волокон цитоскелета или частью сайтов узнавания клетки.
Углеводы — третий важный компонент плазматических мембран. Они всегда находятся на внешней поверхности клеток и связаны либо с белками (образуя гликопротеины), либо с липидами (образуя гликолипиды).Эти углеводные цепи могут состоять из 2–60 моносахаридных единиц и могут быть прямыми или разветвленными. Наряду с периферическими белками углеводы образуют на поверхности клетки специализированные участки, которые позволяют клеткам узнавать друг друга.
Как вирусы заражают определенные органы
Рис. 2. ВИЧ стыкуется и связывается с рецептором CD4, гликопротеином на поверхности Т-клеток, прежде чем проникнуть в клетку или инфицировать ее. (кредит: модификация работы Национального института здравоохранения США / Национального института аллергии и инфекционных заболеваний)
Специфические молекулы гликопротеинов, экспонированные на поверхности клеточных мембран клеток-хозяев, используются многими вирусами для заражения определенных органов.Например, ВИЧ способен проникать через плазматические мембраны определенных видов белых кровяных телец, называемых Т-хелперами и моноцитами, а также через некоторые клетки центральной нервной системы. Вирус гепатита поражает только клетки печени.
Эти вирусы способны проникать в эти клетки, потому что клетки имеют на своей поверхности участки связывания, которые вирусы использовали с одинаково специфическими гликопротеинами в их оболочках. (Фигура 2). Клетка обманывается имитацией молекул вирусной оболочки, и вирус может проникать в клетку.Другие сайты узнавания на поверхности вируса взаимодействуют с иммунной системой человека, побуждая организм вырабатывать антитела. Антитела вырабатываются в ответ на антигены (или белки, связанные с инвазивными патогенами). Эти же сайты служат местами для прикрепления антител и либо уничтожают, либо подавляют активность вируса. К сожалению, эти сайты на ВИЧ кодируются генами, которые быстро меняются, что очень затрудняет производство эффективной вакцины против вируса. Популяция вируса внутри инфицированного человека быстро эволюционирует посредством мутаций в разные популяции или варианты, различающиеся различиями в этих сайтах распознавания.Это быстрое изменение вирусных поверхностных маркеров снижает эффективность иммунной системы человека при атаке вируса, поскольку антитела не распознают новые вариации поверхностных структур.
Фосфолипиды
Как мы только что узнали, основная ткань мембраны состоит из двух слоев молекул фосфолипидов. Гидрофильные или «водолюбивые» области этих молекул (которые выглядят как набор шаров в изображении модели художником) (рис. 1) находятся в контакте с водной жидкостью как внутри, так и снаружи клетки.Таким образом, обе поверхности плазматической мембраны гидрофильны. Напротив, внутренняя часть мембраны между двумя ее поверхностями представляет собой гидрофобную или неполярную область из-за хвостов жирных кислот. Эта область не имеет притяжения для воды или других полярных молекул (мы обсудим это далее на следующей странице).
Гидрофобные или ненавидящие воду молекулы, как правило, неполярны. Они взаимодействуют с другими неполярными молекулами в химических реакциях, но обычно не взаимодействуют с полярными молекулами.При помещении в воду гидрофобные молекулы имеют тенденцию образовывать шар или кластер. Гидрофильные области фосфолипидов имеют тенденцию образовывать водородные связи с водой и другими полярными молекулами как на внешней, так и на внутренней стороне клетки. Таким образом, поверхности мембраны, которые обращены внутрь и снаружи клетки, являются гидрофильными. Напротив, внутренняя часть клеточной мембраны гидрофобна и не взаимодействует с водой. Следовательно, фосфолипиды образуют превосходную двухслойную клеточную мембрану, которая отделяет жидкость внутри клетки от жидкости вне клетки.
Рис. 3. Эта молекула фосфолипида состоит из гидрофильной головки и двух гидрофобных хвостов. Гидрофильная головная группа состоит из фосфатной группы, присоединенной к молекуле глицерина. Гидрофобные хвосты, каждый из которых содержит насыщенную или ненасыщенную жирную кислоту, представляют собой длинные углеводородные цепи.
Молекула фосфолипида (рис. 3) состоит из трехуглеродного глицеринового остова с двумя молекулами жирных кислот, присоединенными к атомам углерода 1 и 2, и фосфатсодержащей группой, присоединенной к третьему атому углерода.
Такое расположение дает всей молекуле область, описываемую как ее голова (фосфатсодержащая группа), которая имеет полярный характер или отрицательный заряд, и область, называемую хвостом (жирные кислоты), которая не имеет заряда. Голова может образовывать водородные связи, а хвост — нет. Молекула с таким расположением положительно или отрицательно заряженной области и незаряженной или неполярной области называется амфифильной или «двоякой».
Эта характеристика жизненно важна для структуры плазматической мембраны, потому что в воде фосфолипиды имеют тенденцию располагаться так, чтобы их гидрофобные хвосты были обращены друг к другу, а их гидрофильные головки были обращены наружу.Таким образом, они образуют липидный бислой — барьер, состоящий из двойного слоя фосфолипидов, который отделяет воду и другие материалы на одной стороне барьера от воды и других материалов на другой стороне. Фактически, фосфолипиды, нагретые в водном растворе, имеют тенденцию спонтанно образовывать маленькие сферы или капли (называемые мицеллами или липосомами), причем их гидрофильные головки образуют внешнюю поверхность, а их гидрофобные хвосты — внутри (рис. 4).
Рис. 4. В водном растворе фосфолипиды имеют тенденцию располагаться так, чтобы их полярные головки были обращены наружу, а их гидрофобные хвосты были обращены внутрь.(кредит: модификация работы Марианы Руис Вильярреаль)
Резюме: Структура клеточной мембраны
Современное понимание плазматической мембраны называется моделью жидкой мозаики. Плазматическая мембрана состоит из бислоя фосфолипидов, причем их гидрофобные хвосты жирных кислот контактируют друг с другом. Ландшафт мембраны усыпан белками, некоторые из которых покрывают мембрану. Некоторые из этих белков служат для транспортировки материалов в клетку или из клетки.Углеводы присоединяются к некоторым белкам и липидам на обращенной наружу поверхности мембраны. Они образуют комплексы, которые служат для идентификации клетки другим клеткам. Жидкая природа мембраны обязана конфигурации хвостов жирных кислот, присутствию холестерина, встроенного в мембрану (в клетках животных), и мозаичной природе белков и комплексов белок-углевод, которые не закреплены прочно в место. Плазматические мембраны ограничивают границы клеток, но они не статичны, а динамичны и постоянно изменяются.
Проверьте свое пониманиеОтветьте на вопрос (ы) ниже, чтобы увидеть, насколько хорошо вы понимаете темы, затронутые в предыдущем разделе. В этом коротком тесте , а не засчитывается для вашей оценки в классе, и вы можете пересдавать его неограниченное количество раз.
Используйте этот тест, чтобы проверить свое понимание и решить, следует ли (1) изучить предыдущий раздел дальше или (2) перейти к следующему разделу.
Клеточная мембрана— подробное руководство
Определение
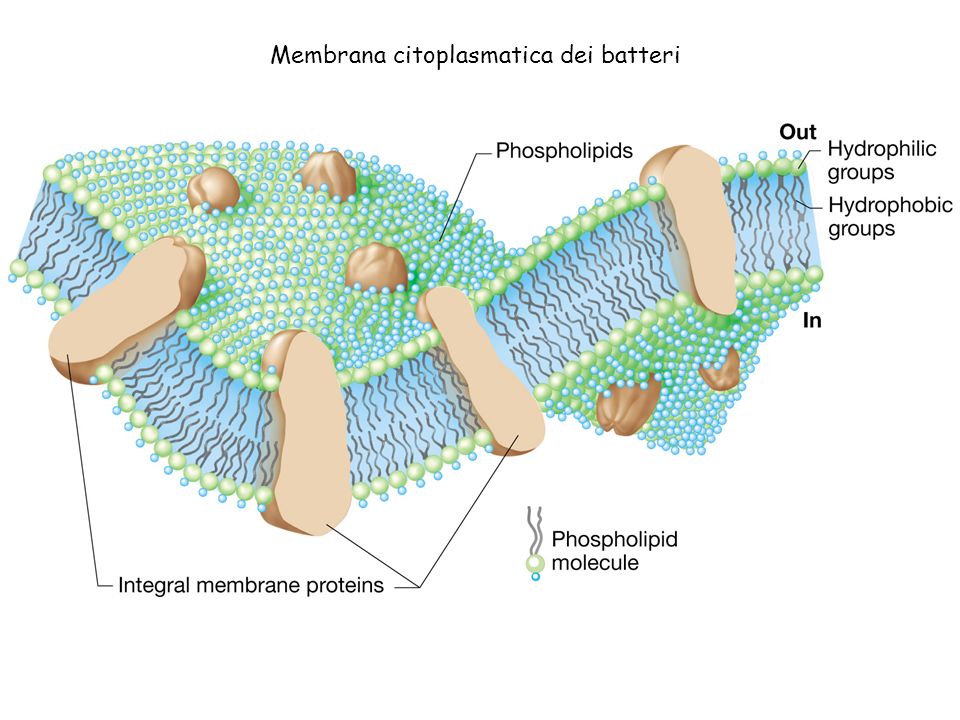
Клеточная мембрана, также известная как плазматическая мембрана, представляет собой двойной слой липидов и белков, окружающий клетку.Он отделяет цитоплазму (содержимое клетки) от внешней среды. Это свойство всех клеток, как прокариотических, так и эукариотических.
трехмерная диаграмма клеточной мембраныФункция клеточной мембраны
Клеточная мембрана придает клетке ее структуру и регулирует материалы, которые входят в клетку и покидают ее. Это избирательно проницаемый барьер, то есть он позволяет одним веществам проникать, но не другим. Подобно подъемному мосту, предназначенному для защиты замка и отпугивания врагов, клеточная мембрана позволяет входить или выходить только определенным молекулам.
Пересекая мембрану
Небольшие молекулы, такие как кислород, который нужен клеткам для выполнения метаболических функций, таких как клеточное дыхание, и двуокись углерода, побочный продукт этих функций, могут легко входить и выходить через мембрану. Вода также может свободно проходить через мембрану, хотя и медленнее.
Однако молекулы с высоким зарядом, такие как ионы, не могут проходить напрямую, как и большие макромолекулы, такие как углеводы или аминокислоты.Вместо этого эти молекулы должны проходить через белки, встроенные в мембрану. Таким образом, клетка может контролировать скорость диффузии этих веществ.
Другой способ, которым клеточная мембрана может переносить молекулы в цитоплазму, — это эндоцитоз. Обратный процесс, когда клетка доставляет содержимое за пределы мембранного барьера, называется экзоцитозом.
Эндоцитоз включает фагоцитоз («поедание клеток») и пиноцитоз («питье клеток»). Во время этих процессов клеточная мембрана образует углубление, окружающее частицу, которую она поглощает.Затем он «отщипывается», образуя небольшую мембранную сферу, называемую пузырьком, которая содержит молекулу и транспортирует ее туда, где она будет использоваться в клетке.
Крупные молекулы могут попадать в клетку в процессе эндоцитоза.Клетки также могут доставлять вещества через клеточную мембрану во внешнюю среду посредством экзоцитоза, который противоположен эндоцитозу. Во время экзоцитоза везикулы образуются в цитоплазме и перемещаются на поверхность клеточной мембраны. Здесь они сливаются с мембраной и выпускают свое содержимое за пределы клетки.Экзоцитоз удаляет продукты жизнедеятельности клетки, которые являются частями молекул, которые не используются клеткой, включая старые органеллы.
Передача сигналов на клеточной мембране
Клеточная мембрана также играет важную роль в передаче сигналов и коммуникации клеток. Мембрана содержит несколько встроенных белков, которые могут связывать молекулы, находящиеся за пределами клетки, и передавать сообщения внутрь клетки.
Важно отметить, что эти рецепторные белки на клеточной мембране могут связываться с веществами, вырабатываемыми другими частями тела, такими как гормоны.Когда молекула связывается со своим рецептором-мишенью на мембране, она инициирует путь передачи сигнала внутри клетки, который передает сигнал соответствующим молекулам.
В результате этих часто сложных сигнальных путей клетка может выполнять действие, указанное сигнальной молекулой, такое как производство или остановка производства определенного белка.
Каким образом структура клеточной мембраны позволяет ей выполнять эти функции?
Структура клеточной мембраны
Фосфолипидный бислой
Клеточная мембрана состоит из фосфолипидного бислоя.Фосфолипиды — это липидные молекулы, состоящие из головы фосфатной группы и двух хвостов жирных кислот. Важно отметить, что свойства молекул фосфолипидов позволяют им самопроизвольно образовывать двухслойную мембрану.
Головка фосфатной группы фосфолипида является гидрофильной, тогда как хвост фосфолипида является гидрофобной. Это означает, что фосфатная группа притягивается к воде, тогда как хвост отталкивается водой.
В воде или водном растворе (в том числе внутри тела) гидрофобные головки фосфолипидов ориентируются так, чтобы находиться внутри, как можно дальше от воды.Напротив, гидрофильные головки будут снаружи, контактируя с водой. В результате образуется двойной слой фосфолипидов, при этом гидрофобные головки сгруппированы вместе в центре, а гидрофильные хвосты образуют внешнюю часть структуры. Технический термин для этого двойного слоя фосфолипидов, который формирует клеточную мембрану, — это бислой фосфолипидов.
Структура клеточной мембраны и связанные с ней компонентыФакторы, ассоциированные с мембраной
Помимо фосфолипидного бислоя, клеточная мембрана также содержит молекулы липидов, особенно гликолипидов и стеринов.Одним из важных стеролов является холестерин, который регулирует текучесть клеточной мембраны в клетках животных. Когда холестерина меньше, мембраны становятся более жидкими, но при этом более проницаемыми для молекул. Количество холестерина в мембране помогает поддерживать ее проницаемость, так что нужное количество молекул может проникать в клетку за раз.
Клеточная мембрана также содержит множество различных белков. Белки составляют около половины клеточной мембраны. Многие из этих белков являются трансмембранными белками, которые встроены в мембрану, но выступают с обеих сторон (т.е.е., они охватывают весь липидный бислой).
Некоторые из этих белков являются рецепторами, которые связываются с сигнальными молекулами. Другие представляют собой ионные каналы, которые являются единственным средством, позволяющим ионам проникать в клетку или выводиться из нее. Ученые используют модель жидкой мозаики для описания структуры клеточной мембраны. Клеточная мембрана имеет жидкую консистенцию из-за того, что большая часть состоит из фосфолипидов, и из-за этого белки свободно перемещаются по ее поверхности. Множество различных белков и липидов в клеточной мембране придают ей мозаичный вид.
Викторина
Библиография
Показать / скрыть
Альбертс, Б., Джонсон, А., Льюис, Дж. И др. Молекулярная биология клетки. 4-е издание. Нью-Йорк: наука о гирляндах; 2002. Липидный бислой. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK26871/
- Lodish, H., Berk, A., Zipursky, S.L., et al. Молекулярная клеточная биология. 4-е издание. Нью-Йорк: У. Х. Фриман; 2000 г.Раздел 3.4, Мембранные белки. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK21570/
- Watson, H. (2015). Биологические мембраны. Очерки биохимии , 59 , 43–69. https://doi.org/10.1042/bse05
Определение мембраны по Merriam-Webster
мем · брана | \ ˈMem-brān \1 : тонкий мягкий гибкий лист или слой, особенно животного или растительного происхождения.
2 : кусок пергамента, составляющий часть рулона
Функция и структура клеточной мембраны
Клеточная мембрана (плазматическая мембрана) — это тонкая полупроницаемая мембрана, окружающая цитоплазму клетки.Его функция заключается в защите целостности внутренней части клетки, позволяя одним веществам проникать в клетку, не допуская попадания других веществ. Он также служит основой для прикрепления цитоскелета у одних организмов и клеточной стенки у других. Таким образом, клеточная мембрана также помогает поддерживать клетку и помогает поддерживать ее форму.
Ключевые выводы
- Клеточная мембрана — это многогранная мембрана, которая окружает цитоплазму клетки. Он защищает целостность клетки, а также поддерживает клетку и помогает поддерживать форму клетки.
- Белки и липиды являются основными компонентами клеточной мембраны. Точная смесь или соотношение белков и липидов может варьироваться в зависимости от функции конкретной клетки.
- Фосфолипиды — важные компоненты клеточных мембран. Они спонтанно образуют липидный бислой, который является полупроницаемым, так что только определенные вещества могут диффундировать через мембрану внутрь клетки.
- Подобно клеточной мембране, некоторые клеточные органеллы окружены мембранами.Ядро и митохондрии — два примера.
Другая функция мембраны — регулировать рост клеток за счет баланса эндоцитоза и экзоцитоза. При эндоцитозе липиды и белки удаляются из клеточной мембраны по мере интернализации веществ. При экзоцитозе везикулы, содержащие липиды и белки, сливаются с клеточной мембраной, увеличивая размер клетки. Клетки животных, клетки растений, прокариотические клетки и клетки грибов имеют плазматические мембраны. Внутренние органеллы также покрыты мембранами.
Липиды клеточной мембраны
Вид фосфолипидов под микроскопом.Stocktrek Images / Getty Images
Фосфолипиды являются основным компонентом клеточных мембран. Фосфолипиды образуют липидный бислой, в котором их гидрофильные (привлеченные водой) участки головы спонтанно располагаются так, чтобы быть обращенными к водному цитозолю и внеклеточной жидкости, в то время как их гидрофобные (отталкиваемые водой) участки хвоста обращены в сторону от цитозоля и внеклеточной жидкости. Липидный бислой является полупроницаемым, что позволяет лишь некоторым молекулам диффундировать через мембрану.
Холестерин — еще один липидный компонент мембран клеток животных. Молекулы холестерина селективно диспергируются между фосфолипидами мембран. Это помогает удерживать клеточные мембраны от жесткости, предотвращая слишком плотную упаковку фосфолипидов. Холестерин не содержится в мембранах растительных клеток.
Гликолипиды расположены на поверхности клеточных мембран и имеют присоединенную к ним углеводную сахарную цепь. Они помогают клетке распознавать другие клетки тела.
Белки клеточной мембраны
Липопротеины и PCSK9 связаны с рецепторами.МАУРИЦИО ДЕ АНДЖЕЛИС / НАУЧНАЯ ФОТОБИБЛИОТЕКА / Getty Images
Клеточная мембрана содержит два типа ассоциированных белков. Белки периферической мембраны находятся вне мембраны и связаны с ней посредством взаимодействия с другими белками. Интегральные мембранные белки вставлены в мембрану и в большинстве своем проходят через мембрану. Части этих трансмембранных белков обнажены с обеих сторон мембраны.Белки клеточной мембраны выполняют ряд различных функций.
Структурные белки помогают придать клеткам поддержку и форму.
Белки рецептора клеточной мембраны помогают клеткам общаться с внешней средой за счет использования гормонов, нейротрансмиттеров и других сигнальных молекул.
Транспортные белки , такие как глобулярные белки, транспортируют молекулы через клеточные мембраны за счет облегченной диффузии.
Гликопротеины имеют присоединенную к ним углеводную цепь.Они встроены в клеточную мембрану и помогают межклеточной коммуникации и транспорту молекул через мембрану.
клеточных мембран извне внутрь — Mellon College of Science
Для клетки мембраны разделяют жизнь на части. Клеточная мембрана, которая окружает всю клетку, физически отделяет внутреннюю часть клетки от внеклеточного пространства. Но это не просто перегородка — клеточная мембрана опосредует транспорт ионов, которые регулируют возбуждение нейронов, обеспечивает место стыковки сигнальных молекул, которые позволяют клеткам общаться друг с другом, и содержит молекулы, которые позволяют иммунной системе идентифицировать клетку как » «я», а не «не-я».«Внутри сложной внутренней части клетки связанные с мембраной компартменты осуществляют важные биохимические процессы, такие как переработка белка и производство энергии.
Несмотря на их важность, еще многое предстоит узнать о мембранах. Поскольку липиды и белки, образующие мембраны, постоянно перемещаются, перемещаются и перестраиваются, чтобы служить потребностям клетки, изучение мембран чрезвычайно затруднено. Этот динамизм ставит ученых в тупик, потому что традиционные экспериментальные методы, такие как кристаллография, не работают с жидкой мембраной, объясняет профессор физики Матиас Лёше.
«Мембраны по своей сути неупорядочены. Если вы хотите что-то изучить с помощью кристаллографии, вам нужно кристаллизовать это, чтобы изучить с помощью рентгеновских лучей. Итак, вам нужна упорядоченная структура, которая повторяется снова и снова. Это то, что вы не можете сделать с мембранами. , поэтому вам необходимо разработать новые методы описания и новые подходы, как в экспериментальной, так и в теоретической физике и биологии, чтобы изучить эти вопросы ».
Ученые из MCS изобрели и используют набор инструментов для более глубокого изучения молекулярных свойств мембран.Эта работа имеет важное значение для понимания нормального поведения клеток и того, что идет не так при таких заболеваниях, как ВИЧ, болезнь Альцгеймера и нарушениях обработки белков, которые приводят к раку и неврологическим расстройствам.
Пробираясь мимо Cellular Sentinel
Наружная мембрана клетки действует как ворота, предотвращая вторжение злоумышленников в клетку. Но вирусам, как и ВИЧ, удается проникнуть через защитный барьер клетки. Стефани Тристрам-Нэгл, доцент исследований в области биологической физики, и ее сотрудник Джон Нэгл, профессор физики и биологических наук, недавно сделали важное открытие, которое помогает понять, почему ВИЧ может с такой очевидной легкостью получать доступ к иммунным клеткам. .
Ученым более 20 лет известно, что ВИЧ сливается с иммунными клетками через gp41, белок, расположенный на поверхности вируса. Хотя у ученых есть рентгеновские изображения gp41 до и после его слияния с клеточной мембраной, до сих пор было загадкой, что именно происходит во время слияния.
Тристрам-Нэгл и Нэгл приготовили стопки из тысяч полностью гидратированных липидных бислоев, используя новый метод, разработанный в их лаборатории. Липиды в нижней части стопки прикреплены к твердой подложке, что придает модельной мембране необходимую стабильность для экспериментального изучения, в то время как липидные бислои наверху стопки сохраняют свою естественную текучесть, что является ключевым требованием для любой биологически значимой модельная система.Тристрам-Нэгл засеял искусственные мембраны гибридным пептидом 23 ВИЧ (FP-23), коротким отрезком gp41, который, как известно, играет ключевую роль в слиянии вирусов. Используя метод диффузного рассеяния рентгеновских лучей, который они впервые использовали, Тристрам-Нэгл и Нэгл количественно определили структурные свойства липидных бислоев в присутствии FP-23. Проанализировав данные диффузного рентгеновского излучения, они обнаружили, что FP-23 резко снижает энергию, необходимую для изгиба мембраны, что значительно упрощает слияние вируса с иммунными клетками и их заражение.
«В клетках мембраны все время изгибаются, что требует энергии», — сказал Тристрам-Нэгл. «Мы обнаружили, что энергия, необходимая для изгиба мембраны, значительно уменьшилась — до 13 раз — когда мы добавили FP-23. Это должно помочь частично объяснить, как легко возникает ВИЧ-инфекция».
Новый поворот в вековой дискуссии
В своей роли привратника клеточная мембрана регулирует молекулярный трафик в клетку и из нее через специализированные мембранные белки. Ионные каналы, белки, покрывающие клеточную мембрану, являются ярким примером.Регулируя ионы, которые входят и выходят из клетки, ионные каналы являются жизненно важным компонентом в инициировании и распространении электрических импульсов в нервных клетках. Дисфункция самого ионного канала или мембраны, в которой он находится, может привести к различным неврологическим расстройствам, включая болезнь Альцгеймера.
В мозгу людей, страдающих болезнью Альцгеймера, имеются нерастворимые бляшки, содержащие неправильно свернутые пептиды, называемые бета-амилоидом (Aβ). Бляшки Aβ накапливаются между нервными клетками и участвуют в заболевании с тех пор, как Алоис Альцгеймер впервые открыл их 100 лет назад.В последние годы ученые предположили, что олигомеры Aβ — агрегаты Aβ, промежуточного звена между единичным пептидом и зрелыми бляшками — каким-то образом взаимодействуют с мембранами нервных клеток, но фактический механизм токсичности клеток остается неясным.
«Мы знаем, что олигомеры Aβ сильно взаимодействуют с мембранами и препятствуют их способности сохранять ионные градиенты между внутренней и внешней частью клетки. Однако остается огромной проблемой определить, действительно ли олигомеры Aβ делают отверстие в мембране, или если они влияют на свойства мембраны настолько, чтобы изменить критические свойства мембранных каналов », — объясняет Лёше.
Используя методы рассеяния нейтронов, Лёше и его коллеги из Национального института стандартов и технологий (NIST) изучают, как Aβ взаимодействует с моделями синтетических мембран, называемыми «привязанными двухслойными липидными мембранами» (tBLM). TBLM состоят из липидного бислоя, который химически связан с твердой подложкой, например с кремниевой пластиной, через полимерные тросы.
«Эти привязанные мембраны очень мощные, потому что они чрезвычайно стабильны. Мы можем манипулировать ими и измерять их в течение длительного времени.»Это необычно для изысканно хрупкого листочка жидкого материала толщиной 5 нанометров», — сказал Лёше.
В сотрудничестве с химиками из Калифорнийского университета в Ирвине команда Лёше инкубировала tBLM с олигомерами Aβ и изучила структурную и функциональную реакцию мембраны в Центре нейтронных исследований NIST в Гейтерсбурге, штат Мэриленд. Они наблюдали нарушение изоляционных свойств липидный бислой, который вызывает утечку ионов через мембрану. Но характер утечки мембраны отличается от сигнатуры других мембранных дисфункций, которые группа Лёше подробно изучила.Например, некоторые бактерии выделяют токсин, альфа-гемолизин, который внедряется в мембраны клетки-хозяина, образуя каналы, которые вызывают утечку заполненных водой ионов. По словам Лёше, при сравнении режима работы альфа-гемолизина и Aβ становится ясно, что олигомеры Aβ не просто «пробивают дыры» в мембране.
Группа Лёше теперь расширяет эту работу в сотрудничестве с Маркусом Дезерно, доцентом физики, который разрабатывает компьютерные модели клеточных мембран.
«Вы можете сделать так много с помощью экспериментов, но из-за внутреннего расстройства биологической мембраны невозможно рассмотреть все аспекты атомных деталей или молекулярной динамики.Все это можно сделать на компьютере », — объясняет Дезерно.
Дезерно и его коллеги из Института исследований полимеров Макса Планка в Майнце, Германия, создали компьютерное моделирование, которое следует за поведением искусственной мембраны, состоящей из 50 000 отдельных молекул липидов. Каждая молекула липида просто представлена в виде трех сфер.
«Наша модель крупнозернистая, — поясняет Дезерно. «Вы можете думать об этом как о картине импрессионизма. На расстоянии все выглядит хорошо.Вы можете увидеть водяные лилии или балерин. Но вблизи все детали исчезли; вы просто видите цветные пятна. Мы заинтересованы в том, что происходит с кувшинками, а не в цветных пятнах », — говорит он.
С помощью этой крупнозернистой модели Дезерно может уловить важные характеристики, такие как изгибы и изгибы мембраны, что позволяет ему задавать вопросы, выходящие за пределы атомарного уровня, но меньшие, чем уровень всей клетки. Его модель также универсальна; он может добавить определенные представляющие интерес белки к липидной мембране и наблюдать, как они взаимодействуют.Следующий шаг для Дезерно и Лёше — засеять искусственную мембрану в компьютерной модели Дезерно белками Aβ, чтобы получить больше подсказок о том, как Aβ повреждает мембрану.
с мембраной
Наружная мембрана клетки — не единственная мембрана, выполняющая важные жизненно важные процессы. Клетки делят свои внутренние части на связанные с мембраной органеллы, такие как эндоплазматический ретикулум (ЭР) и аппарат Гольджи, для более эффективного выполнения различных задач, таких как производство белка.
Но мембрана органелл — это гораздо больше, чем просто барьер. Мембраны играют ключевую роль в транспортировке белков из ER в аппарат Гольджи, внутри аппарата Гольджи, а затем из Гольджи к их конечному месту назначения в клетке.
«Многие болезни возникают из-за нарушения в работе мембран, — сказал Адам Линстедт, профессор биологических наук. Исследователи обнаружили, что десятки генетических нарушений у человека являются результатом дефектов мембранного обмена, включая несколько нейродегенеративных заболеваний и нарушений развития.
Линстедт вместе с Кристиной Ли, доцентом биологических наук, исследуют пути мембранного переноса в Гольджи и ER и многое узнают о структуре органелл на этом пути.
Линштедт идентифицировал группу белков, участвующих в формировании ленты Гольджи, сложной структуры субкомпартментов Гольджи или стопок, связанных между собой канальцами. Стеки и ферменты внутри них функционируют как сборочная линия, обрабатывая тысячи вновь синтезированных белков и липидов, проходящих через Гольджи.Когда недавно синтезированный белок перемещается в стеке, ферменты модифицируют белок, добавляя такие компоненты, как углеводы или фосфаты. К тому времени, как белок покидает Гольджи, он полностью обработан. Используя метод, называемый интерференцией РНК, Линстедт подавил экспрессию белков Гольджи GM130 и GRASP65 и обнаружил, что стеки Гольджи не соединяются в ленту. В клетках без ленты Линстедт обнаружил, что некоторые из несвязанных стопок имеют более высокие уровни ферментов, а другие — более низкие, в отличие от случаев, когда стопки ориентированы в виде ленты и имеют равное распределение ферментов Гольджи.Согласно Линстедту, единообразные уровни ферментов могут иметь решающее значение, потому что клетки с несвязанной лентой Гольджи имеют недостаточно обработанные белки. Нарушение обработки может привести к серьезным дефектам развития у множества организмов, от мышей до людей.
«Никто не знал, для чего нужна лента», — объясняет Линстедт. «Теперь у нас есть одно объяснение — лента важна для уравновешивания концентрации фермента во всей мембранной сети и необходима для правильной обработки белков.«
Гольджи играет важную роль в окончательной обработке белков, но изначально белки собираются в ER, единой непрерывной мембранной сети, которая простирается от ядра до клеточной мембраны. Ли применяет биохимический подход к изучению ER, разрушая открытые клетки и промывая солью, которая удаляет молекулы, электростатически связанные с мембраной ER. После тестирования этих молекул по одной, чтобы увидеть их влияние на мембрану ER, Ли определил ключевой фактор, участвующий в формировании расширенной сети ER.Фактор, вариант фермента нуклеозиддифосфаткиназы (NDKB), уже был известен своей функцией в клетках, но не был связан с морфологией мембраны. Оказывается, что NDKB связывается непосредственно с кислыми фосфолипидами в мембране ER и может собираться с образованием каркаса, который стабилизирует расширенную мембранную сеть.
«Существует огромный поток мембраны, который начинается в ER и движется к поверхности клетки», — объясняет Линстедт. «Большая часть этого мембранного потока создается везикулами, образующимися из одного отсека и сливающимися со следующим отсеком.Возможно, если мы лучше поймем эти процессы, мы сможем нацелить мембранные пути трафика для борьбы с болезнями ».
По мере того, как ученые MCS используют инструменты биологии, чтобы понять основные функции клеток, и инструменты физики, чтобы выявить физические свойства мембран, они продвигаются на один шаг ближе к пониманию того, как мембраны функционируют при здоровье и болезнях.
«Я считаю, что если биологи, физики, инженеры и математики вместе будут ломать голову, мы придем к новым открытиям намного быстрее и с большей уверенностью», — сказал Дезерно.
мембран | SEER Обучение
Мембраны тела — это тонкие слои ткани, которые покрывают тело, выстилают полости тела и покрывают органы внутри полостей полых органов. Их можно разделить на эпителиальные и соединительнотканные мембраны.
Эпителиальные мембраны
Эпителиальные мембраны состоят из эпителиальной ткани и соединительной ткани, к которой она прикреплена.Двумя основными типами эпителиальных оболочек являются слизистые оболочки и серозные оболочки.
Слизистые оболочки
Слизистые оболочки — это эпителиальные оболочки, которые состоят из эпителиальной ткани, прикрепленной к подлежащей рыхлой соединительной ткани. Эти оболочки, иногда называемые слизистыми оболочками, выстилают полости тела, которые открываются наружу. Весь пищеварительный тракт выстлан слизистыми оболочками. Другие примеры включают дыхательные, выделительные и репродуктивные пути.
Серозные мембраны
Серозные оболочки выстилают полости тела, которые не открываются непосредственно наружу, и покрывают органы, расположенные в этих полостях. Серозные оболочки покрыты тонким слоем серозной жидкости, секретируемой эпителием. Серозная жидкость смазывает мембрану и уменьшает трение и истирание, когда органы грудной или брюшно-тазовой полости движутся друг относительно друга или стенки полости. Серозным оболочкам даны специальные названия в зависимости от их расположения.Например, серозная оболочка, выстилающая грудную полость и покрывающая легкие, называется плеврой.
Мембраны соединительной ткани
Соединительнотканные оболочки содержат только соединительную ткань. Синовиальные оболочки и мозговые оболочки относятся к этой категории.
Синовиальные мембраны
Синовиальные мембраны — это соединительнотканные мембраны, выстилающие полости подвижных суставов, таких как плечо, локоть и колено. Подобно серозным оболочкам, они выстилают полости, которые не открываются наружу.В отличие от серозных оболочек они не имеют слоя эпителия. Синовиальные оболочки выделяют синовиальную жидкость в полость сустава, и это смазывает хрящи на концах костей, чтобы они могли двигаться свободно и без трения.